Клеточный уровень.
ВИЧ поражает различные клетки человека, но два типа являются критическими для развития ВИЧ-инфекции — это Т-лим- фоциты-хелперы и макрофаги. Если первые постепенно устраняются из крови самим вирусом, то вторые, наоборот, приобретают устойчивость к апоптозу и используются им до самой гибели больного СПИДом.
Еще в 1994 г. М. Stevenson и Н. Е. Gendelman на основе собственных наблюдений пришли к выводу, что макрофаги играют центральную роль в прогрессировании ВИЧ-инфекции и ее клинических проявлений. Вирус приникает в организм человека как в составе макрофагов, так и с тканевыми жидкостями в виде свободных частиц. Макрофаги распространяют ВИЧ по тканям и органам и непосредственно передают его Т-клеткам-хелперам. В этом процессе Т-хелперы активируются через представление антигена главным комплексом гистосовместимости макрофагов. Вирус от моноцитов передается к взаимодействующим с ними Т-хелперами. Активация макрофагов происходит через опортунистическую инфекцию и стимуляцию иммунной системы (рис. 56).Поведение макрофагов, вышедших из под контроля T- и В-клеточной системы, мало похоже на то, что приписывается им учебниками по иммунологии. По «законченным» представлениям их авторов о роли иммунной системы в инфекционном процессе, макрофаги относятся к врожденной системе иммунитета человека. Основное предназначение макрофага — удаление из крови микроорганизмов, опухолевых клеток, иммунных комплексов, презентация антигенов лимфоцитам и т. п. (см., например, работы Кузника Б. И., 2004; Ройта А. С. с соавт., 2000; Галактионова В. Г., 1998), В раза. 2.2 уже обращено внимание исследователей на то, что ВИЧ взаимодействует с макрофагами не только посредством хемокиновых рецепторов,
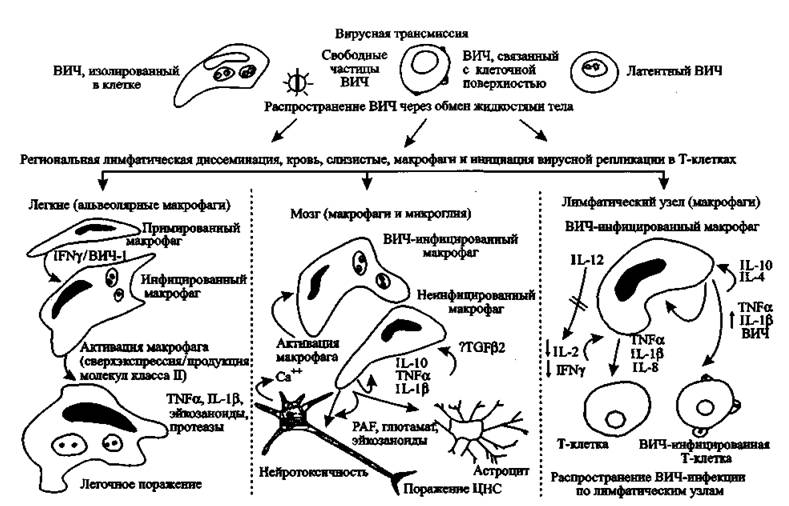
Рис. 56. Родь макрофагов в прогрессировании ВИЧ-инфекции по М. Stevenson, И.
Е. Gendelman (І994). Современную модель повреждения и смерти нейронов под воздействием ВИЧ-инфекции см. на рис. 34но и маннозных, что характерно для взаимоотношений паразитов почвенных и водных простейших. В последнем случае он ведет себя как эндосимбионт, т. е. не интегрируется с геномом макрофага и не реплицируется. Хемокиновые рецепторы на поверхности макрофагов, с которыми взаимодействует ВИЧ, очень древние, так как принадлежат хемокинам C-X-C и C-C подклассов.
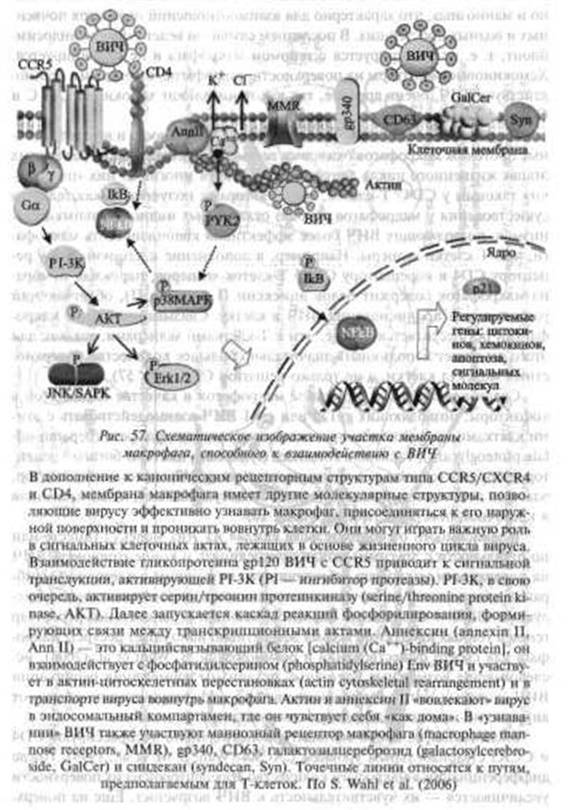
По данным S. Walil et аі. (2006), клеточная поверхность и внутриклеточные протеины макрофагов оказались весьма эффективными на различных этапах жизненного цикла лентивирусов. И хотя многие из них «подражают» таковым у CD4+ Т-клеток, этими авторами получены доказательства существования у макрофагов хорошо отлаженных индивидуальных механизмов, позволяющих ВИЧ более эффективно колонизировать макрофаги, чем Т-клетки-хелперы. Например, в дополнение к хемокиновому рецептору СІМ и корецептору CCR5 Т-клеток-хелперов, наружная мембрана макрофагов содержит белок аннексии II (annexin II), облегчающий ранние этапы проникновения ВИЧ в клетку. Связывание ВИЧ с макрофагами осуществляется проще, чем с Т-клетками-хелперами, так как для этого он может использовать значительно большее количество поверхностных молекул клетки, а не только рецептор CD4 (рис. 57).
Среди компонентов мембраны макрофагов в качестве кандидатов в кофакторы, позволяющих gpl20 или gp41 ВИЧ взаимодействовать с этими клетками, уже описаны протеогликаны гепарин-сульфата (heparan sulfate proteoglycans), такие как синдекан (syndecan); цистеин-богатый рецептор-мусорщик (cysteine-rich scavenger receptor), gp340; маннозный рецептор, эластаза лейкоцитов человека (human leukocyte elastase), локализованная в клеточной поверхности и др. (табл. 16).
В зависимости от обстоятельств каждая из этих молекул (вместе или по отдельности с другими) может присоединять и концентрировать ВИЧ на поверхности макрофага, обеспечивать ему слияние с клеточной мембраной и/или влиять на его «благополучие» посредством сигнальной транс- дукции.
ВИЧ — облигатный паразит, имеющий ограниченный репертуар генов и генных продуктов, весьма эффективно использует ресурсы макрофага для своего существования и размножения, S. Wahl et аі. (2006), исследовавшие возможные клеточные механизмы торможения репликации ВИЧ, которые могли бы рассматриваться как «защищающие клетку от вируса», не нашли убедительных доказательств их наличия.Выделенные из крови моноциты конститутивно экспрессируют CD4 и CCR5 с вариабельным уровнем CXCR4 (Wahl S. M et аі., 1999). После дифференциации в макрофаги количество корецепторов на их поверхности увеличивается — их чувствительность к ВИЧ возрастает. Еще на поверх-

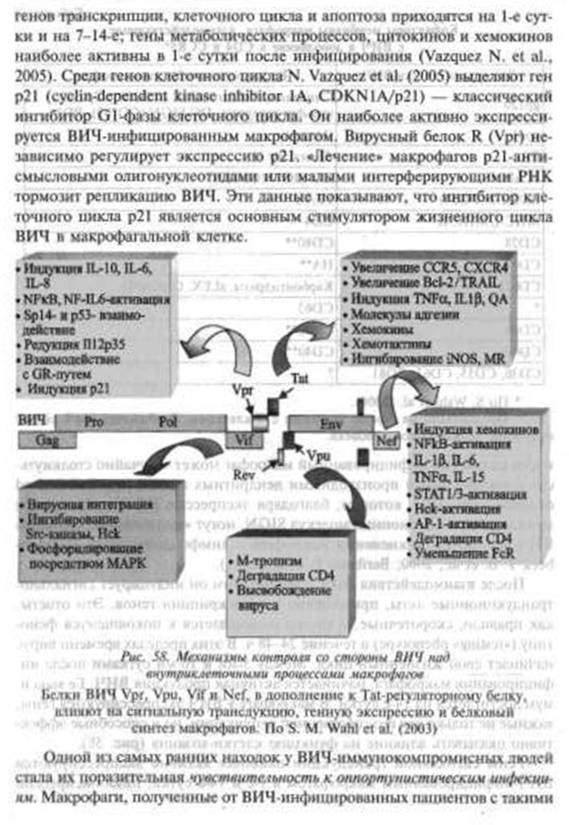

Уровень генома. Предположения о связи между собой процессов, в κοτορίίχ участвуют экзогенные ретровирусы и эндогенные ретроэлементы, появились в начале 1990-х гг. Однако трудности их изучения обусловлены не только недостаточным развитием соответствующей методологии. В привычном для человека масштабе времени не всегда возможно определение пределов таких процессов в эволюционной истории его как вида Homo sapiens. Пока «максимальным разрешением* таких исследований является жизнь конкретного человека. В этом масштабе времени нециклические процессы, в основе которых лежит взаимодействие эндогенных ретровирусов и ретроэлементов с экзогенными ретровирусами, могут, как минимум, протекать в следующих четырех вариантах.
1. Эндогенные ретровирусы и ретроэлементы усиливают инфекционный процесс, вызванный экзогенным ретровирусом.
Н. В. Umovitz и W1 Н. Murphy(1996) описали ВИЧ-инфицированного пациента, чья иммунная система, благодаря активности эндогенного ретровируса млекопитающих типа С, находилась в хронически активированном состоянии. В результате прогрессирование ВИЧ-инфекции шло очень стремительно.
2. Эндогенные ретровирусы и ретроэлементы участвуют в комплементации нарушенных функций экзогенных ретровирусов. Отдельные HERV-K имеют транскрипционно активные открытые рамки считывания и кодируют собственную протеазу, идентичную протеазе ВИЧ. Протеаза HERV-K может комплементировать функцию протеазы ВИЧ у ВИЧ-инфицированных пациентов, подвергнутых лечению ингибиторами протеаз, и тем самым значительно снизить эффективность таких препаратов (Padow М, et аі., 2000).
3. Экзогенные ретровирусы активизируют эндогенные ретровирусы и ретроэлементы. J. J. Goedert et аі. (1999) показали усиление экспрессии генов эндогеного ретровируса KlO (HERV-KlO) у ВИЧ-инфицированных людей и больных СПИДом и, соответственно, повышение риска развития у них тестикулярного рака (testicular cancer). ВИЧ индуцирует появление вирусных частиц HERV-K(HML-2) в сыворотке крови человека (Contreras- Galindo R. et аі., 2006; 2007).
По данным R. Contreras-Galindo et аі. (2007), РНК HERV-K(HML-2) встречается в сыворотке крови ВИЧ-инфицированных пациентов почти всегда, в 95,33 % исследуемых образов, но отсутствует в крови неинфи- цированных ВИЧ людей. Ранее Contreras-Galindo R. et аі. (2006), исследовав на нагрузку по РНК HERV-K кровь двадцати ВИЧ-инфицированных пациентов, принимавших высокоактивную ретровирусную терапию (HAART), обнаружили, что титры такой РНК всегда значительно повышены у пациентов с несупрессированной HAART (non-suppressive HAART), т, е. у тех из них, у которых такая терапия не дает эффекта в субоптимальных дозах. У пациентов, у которых HAART давала положительный эффект, повышения титров РНК HERV-K не обнаруживалось. В описанных случаях имеет место активизация ВИЧ HERV-K, которые, в свою очередь, снижают эффективность HAART.
В качестве активаторов HERV могут выступить не только экзогенные ретровирусы, например, ВИЧ. По крайней мере, в клеточных линиях, инфицированных вирусом гриппа A/WSN/33 или вирусом герпеса первого типа, удавалось достичь аберрантной экспрессии HERV семейства W (Nellaker С. et аі,, 2006).
Пока неизвестно, способны ли передаваться горизонтально между людьми HERV, ативизировавшиеся благодаря ВИЧ или другой инфекции. Но такую возможность нельзя исключать, и, несомненно, это очень интересная тема будущих исследований взаимоотношений экзогенных и эндогенных ретровирусов человека.
4. Эндогенные ретровирусы и ретроэлементы тормозят инфекционный процесс, вызванный экзогенным ретровирусом. Экспрессия Fvl, gag-последовательности эндогенных ретровирусов мышей, гомологичных семейству HERV-L, блокирует отдельные штаммы вируса лейкемии мышей. Каким образом это происходит, неясно (Bannert N., Kurth R., 2004),
Патологические состояния, в основе которых лежит автономная деятельность эндогенных ретровирусов и ретроэлементов, описаны в разд, 4,4.
Ф* Ф
При ВИЧ-инфекции клеточная и гуморальная составляющие иммунной системы постепенно перестают контролировать реликтовую иммунную систему, и фагоцитирующие клетки начинают играть в инфекционном процессе ту же роль «мусорщиков», которую они играли у первых многоклеточных животных. Между такими клетками и их паразитами восстанавливаются связи, отношения и даже иерархия, существовавшие у них в архее. На уровне макроорганизма они проявляются десятками нециклических инфекционных процессов, называемых СПИД-ассоциируемыми, и активизацией эндогенных ретроэлементов.
Так как взаимодействие возбудителей СПИД-ассоциируемых инфекций, ВИЧ и клеток иммунной системы носит специфический характер на надклеточном, клеточном и генетическом уровнях, я предлагаю выделить такие процессы в отдельную группу — многокомпонентные нециклические инфекционные процессы. Их сложность, по мере ослабления контроля клеточной и гуморальной иммунной системы над реликтовой иммунной системой, нарастает; и они начинают поддерживать себя сами по различным дублируюшим специфическим механизмам.
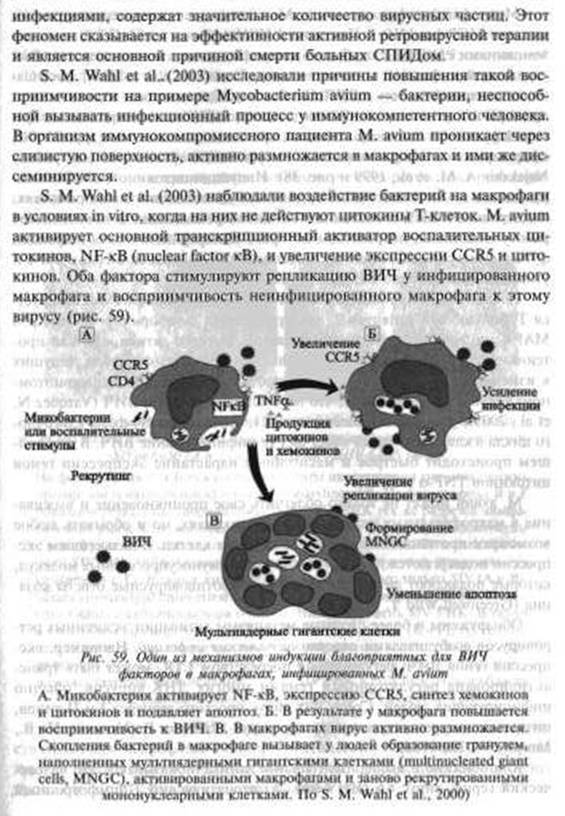
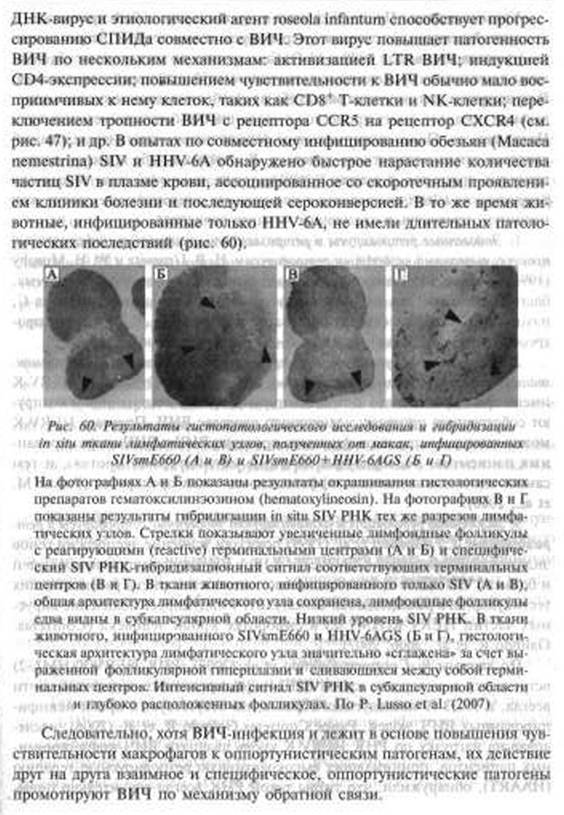
Например, белок SP-A, присутствующий в бронхоальвеолярной жидкости ВИЧ-инфицированных людей, усиливает прикрепление М. tuberculosis к альвеолярным макрофагам; CDKNlA/p21 — классический ингибитор Gl-фазы клеточного цикла индуцируется как ВИЧ, так и микобактериями, но одновременно он является стимулятором жизненного цикла и ВИЧ, и микобактерий в макрофагальной клетке; индукция М. avium основного транскрипционного активатора воспалительных цитокинов, NF-κΒ, ведет к увеличению экспрессии CCR5 и цитокинов, стимулирующих репликацию ВИЧ; экспрессия генов ВИЧ, регулируемая посредством LTR, может быть трансакти- вирована регуляторными генами многих ДНК-вирусов; они же способны повысить чувствительность к ВИЧ у CDg+ Т-клеток и NK-клеток; «переключить» тропность ВИЧ с корецептора CCR5 на корецептор CXCR4. После утраты иммунной системой человека T- и В-клеточной функции ВИЧ переходит в разряд «равного среди равных» формирующегося паразитоценоза первых многоклеточных организмов. Отдельные нециклические инфекции могут развиваться как комплекс инфекционных болезней и болезней, традиционно не относимых к инфекционным (онкологические, аутоиммунные и др.).
Еще по теме Клеточный уровень.:
- окислительный стресс и повышенный уровень ПОЛ при старении животных
- 2.5.2. Повышенный уровень ПОЛ при лейкозогенезе
- клеточная пролиферация
- Роль белков теплового шока в Т-клеточной регуляции хронического воспаления
- Молекулярный шаперон HSP70 и скорость клеточной пролиферации в онкогенезе и риске развития метастазов при инвазивном раке молочной железы
- 2.1. Исследование экспрессии мРНК гена вЗА-адаптина в опухолях шейки матки и в клеточных линиях рака шейки матки.
- Клеточный уровень.
- КЛЕТОЧНЫЕ ЭФФЕКТОРЫ И МЕДИАТОРЫ ВОСПАЛЕНИЯ
- КАНЦЕРОГЕНЕЗ И ФАКТОРЫ КЛЕТОЧНОГО РОСТА
- Анализ кинетики накопления 5-АЛК-индуцированного Пп IX в аутоидентичных клеточных культурах
- КЛЕТОЧНЫЙ И ТКАНЕВОЙ УРОВНИ АДАПТАЦИЙ У ПОЙКИЛОТЕРМНЫХ ОРГАНИЗМОВ
- КЛЕТОЧНЫЙ И ТКАНЕВОЙ УРОВНИ АДАПТАЦИЙ У ГОМОЙОТЕРМНИХ ОРГАНИЗМОВ
- Специфические свойства клеточных культур
- Уровни регуляции клеточного ответа.
- Тема № 6. Белки - маркеры клеточного цикла
- Тема № 8. Белковые молекулы, характеризующие клеточную адгезию
- 3.3 Действие Rpf на клеточную стенку Л/, tuteus
- 6.1.Обоснование возможности использования инкубации клеточной массы крови с антибиотиками для лечения больных с ГВЗПМ
- Изучение состояния системного клеточного иммунитета у часто болеющих детей с патологией ЛОР-органов.