клеточная пролиферация
является непременной предпосылкой возникновения неоплазмы. Без пролиферации клеток ни один канцероген не способен индуцировать опухоли, т.е. клеточная пролиферация необходима для канцерогенеза, но всё же недостаточна в качестве «канцерогенного фактора» в стандартном канцерогенном тесте (Zito, 1992).
Приобретение способности к пролиферации определённой адаптировавшейся частью клеток при хроническом воспалении означает, что вероятность опухолевой трансформации этих клеток в случае появления других необходимых условий значительно увеличивается. В то же время адаптировавшаяся часть воспалённой ткани «подготовлена» к злокачественному перерождению и по другому признаку – поддержанию в её клетках повышенного уровня ПОЛ, который, согласно кис-лородно-перекисной модели канцерогенеза, может быть непосредственно причастным к указанной трансформации на последующих этапах хронического воспалительного процесса.Если же иметь в виду острое воспаление, то слишком высокий уровень пер-оксигеназных процессов лишает возможность адаптации к ним даже малому числу клеток в области воспаления, и все они подвергаются апоптозу или окислительному разрушению. В этом видится одна из причин того, что острый вос-палительный процесс не только не способствует развитию опухоли, но и подавляет его. Здесь как раз тот случай, когда внутриклеточный дисбаланс про- и антиоксидантных процессов возрастает до значений, соответствующих апоптоз-ному ∆А2 (ПО – АО) или цитолизному ∆Ц (ПО – АО) диапазонам (см. п. 7.1).
Таким образом, по излагаемому здесь представлению длительно протекающие воспаления в силу окислительного стресса и возбуждаемой в части клеток пролиферации создают для перерождения единичных клеток этой категории необходимые с точки зрения указанной модели условия. В таком именно качестве хроническое воспаление может рассматриваться как пре- и пронеопластическая стадия, которая постепенно перерастает в настоящий канцерогенез, реализуемый по кислородно-перекисному механизму.
Со сказанным согласуются данные о том, что in vitro генерируемые воспалительными фагоцитами оксиданты могут выступать как полные канцерогены, т.е. вызывать как инициацию, так и промоцию. Однако in vivo, как считают (Weitzman,Gordon, 1990), эти метаболиты функционируют как опухолевые промоторы или коканцеро-гены из-за высоких уровней эндогенной антиоксидантной защиты. Это мнение, на наш взгляд, находится в противоречии с индуцированием in vivo неоплазм инородными химически инертными телами – процессом, в ходе которого макрофаги атакуют чужеродное вещество, но неизбежно создают и поддерживают полноценную канцерогенную окислительно-воспалительную среду в прилежащих к ним слоях клеток (см. главу 5).В пользу изложенных выше взглядов на общность пероксигеназных процессов при воспалении и канцерогенезе свидетельствует наличие эндогенных и экзогенных соединений, обладающих одновременно противовоспалительными и противоопухолевым действием. Нами, в частности, уже упоминалось профилактическое и лечебное действие SOD и церулоплазмина при воспалении. Но эти же вещества известны и как антинеопластические. Так, церулоплазмин тормозит рост различных экспериментальных опухолей на 25-50 % (Бердинских и др., 1984). Как и в случае воспалений, концентрация церулоплазмина у боль-ных злокачественными опухолями значительно выше, чем у здоровых людей и при доброкачественных новообразованиях (Vlad et al., 1998). Подобная реакция опухолевого организма подтверждает способность его к мобилизации различных антиоксидантных средств (Лю, Ефимов, 1976).
Двойственный эффект проявляет также соединение меди – Cu(3,5-диизо-пропилсалицилат)2, обладающий активностью SOD, но значительно лучше её проникающий в клетку. Подобно указанному соединению действуют и фенольные антиоксиданты (Kensler et al., 1983). Инициирующим и активирующим канцерогенез агентам могут противодействовать антивоспалительные стероиды и ретиноидная кислота. Общим для всех указанных противовоспалительных и противоопухолевых воздействий является их антиоксидантная и, в частности, антисупероксидная направленность.
* * *
Распространённое мнение о том, что фактическая гипоксия характерна для всех опухолевых клеток и даже служит необходимым фоном при развитии новообразований (Мосиенко и др., 1985), представляется спорным или просто не логичным.
До сих пор в отдельных публикациях с условием гипоксии связывают прогрессирование опухолевого процесса, усиление метастазирования и, вообще, бoльшую агрессивность опухоли и худшие прогнозы (см. обзор: Beinert et al., 1999). Внутриклеточная гипоксия и признаваемое многими исследователями усиление свободнорадикальных процессов ПОЛ in vivo – события в принципе одновременно возможные (Кожевников, 1985), но, с нашей точки зрения, маловероятные. К выводу об активации ПОЛ при гипоксии обычно приходят на основании экспериментов с ишемизированными органами. В действительности же такое представление является ошибочным, поскольку последующая часть эксперимента (выделение и исследование внутриклеточного материала из ишемизированных тканей и органов) традиционно проводится в гипероксической насыщенной воздухом среде (см. главу 4, п. 4.2.4). Избыточное ПОЛ в мембранах естественнее связать с гипероксией, которая постулируется нами в клетках активно растущих участков неоплазмы, а также при первичном возникновении таких клеток. Гипероксия есть следствие прямого или косвенного нарушения различными канцерогенными факторами (химическими, радиационными и др.) митохондриального дыхания, ведущего к состоянию лишь фиктивной гипоксии. В такой постановке первопричина гипероксии и пероксигенации при опухолевой трансформации до наших первых сообщений (Лю, Ефимов, 1976; Лю, Саприн, 1980) в литературе не рассматривалась, не было и системного подхода к объяснению с позиций предложенной концепции множества разнородных, разобщённых и даже, на первый взгляд, противоречивых фактов.О том, что факторы, влияющие на эффективность действия дыхательной цепи митохондрий – основного потребителя О2 в клетке, могут определять канцерогенную ситуацию в ней показывает работа, в которой анализировали экспрессию гена цитохром-с-оксидазы при дифференцировке и трансформации клеток толстой кишки человека, взятых из биопсийных образцов нормальных и злокачественных тканей. Экспрессия указанного гена, как оказалось, связана с нормальной дифференцировкой эпителиальных клеток толстой кишки, а сни-жение экспрессии может быть маркёром увеличения степени риска развития рака толстой кишки, причём указанные сдвиги в экспрессии не обусловлены изменением числа геномов митохондрий (Heerdt et al., 1990).
В принципиаль-ном плане эти данные могут означать, что в клетке существует отдельная система регуляции интенсивности дыхания с отрицательной обратной связью, действующая путём изменения уровня экспрессии гена терминального фер-мента дыхательной цепи. Настройка этой системы в норме может, вероятно, корректироваться внеклеточными сигналами применительно к различным эта-пам онтогенеза, а какие-либо нарушения в ней могут быть ответственными за дефицит цитохромоксидазы и соответственно за внутриклеточную гипероксию и избыточную пероксигенацию, опасных в канцерогенном отношении.По тем же соображениям любое соединение, способствующее образованию избыточной концентрации свободных радикалов О2, в принципе канцерогенно или «работает» на канцерогенез. Участие этих радикалов и продуктов липидной пероксигенации в разных этапах канцерогенеза считается доказанным, а недостаточность антиоксидантной системы клетки (ферментативной и неферментативной её составляющих) – тесно связанной с повышением риска развития рака у человека (Krämer, 1994; Rautalahti, Huttunen, 1994 и др.) и, более того, необходимым условием для возникновения и протекания этой болезни (Лю, Шайхутдинов, 1991). Поток научной информации в пользу этих положений непрерывно нарастает. С данной логикой коррелируют и материалы следу-ющего обобщения (Zatterale et al., 1999): многие наследственные болезни (синдромы с хромосомной нестабильностью, пигментная ксеродерма, синдромы Вернера, Блума, Дауна, атаксия-телеангиэктазия и др.) предрасположены к раку прежде всего потому, что их патогенетический механизм связан с окси-дативным стрессом.
Многочисленные факты подтверждают важность создания в клетке усло-вий повышенного свободнорадикального окисления различных субстратов для перерождения её в опухолевую, развития из неё очага новообразования. Онкогенная роль АФК подтверждается и канцерогенностью озона (Victorin, 1996) – сильнейшего окислителя, образующего при действии на органические и неорганические вещества цитотоксические перекисные соединения.
По существу же, к генерации О , ОН˙ и Н2О2 прямо или косвенно причастны самые различные экзогенные и эндогенные агенты и факторы, в том числе некоторые антибиотики, металлы и иммунокомпетентные клетки; пролифераторы пероксисом, различные виды излучений, сигаретный дым; ингибиторы антиоксидантных и дыхательных ферментов, инородные трудноокисляемые тела в тканях и многие другие (Лю, Шайхутдинов, 1991).
, ОН˙ и Н2О2 прямо или косвенно причастны самые различные экзогенные и эндогенные агенты и факторы, в том числе некоторые антибиотики, металлы и иммунокомпетентные клетки; пролифераторы пероксисом, различные виды излучений, сигаретный дым; ингибиторы антиоксидантных и дыхательных ферментов, инородные трудноокисляемые тела в тканях и многие другие (Лю, Шайхутдинов, 1991).Давно уже известная эмпирически установленная зависимость эффективности канцерогенеза от размножения клеток до сих пор не имеет убедительного научного обоснования. В самом деле, почему бoльшая часть опухолей воз-никает в органах и тканях с обновляющимся клеточным составом и довольно высоким уровнем пролиферации? И почему предпочтительным объектом малигнизации, как считают (Урываева, 1990), являются недифференцированные клетки стволовых подразделений ткани? Если исходить из положений изложенной нами концепции, то основным и наиболее принципиальным видится утверждение: в нормальных ростстимулированных клетках умеренное возрастание уровня свободнорадикальных перекисных процессов необходимо для реализации окислительного митогенеза, но это же условие в более выраженном варианте лежит и в основе кислородно-перекисного механизма канцерогенеза. В указанном смысле нормальные размножающиеся клетки физиологически уже «подготовлены и приближены» природой к опухолевой трансформации. При воздействии факторов, способных устойчиво перевести дисбаланс ∆ (ПО – АО) с «пролиферативного» уровня ∆П (ПО – АО) на более высокий, «канцерогенный» ∆К (ПО – АО), эти клетки могут сравнительно легко (по сравнению с непролиферирующими дифференцированными) переродиться в опухолевые.
Действительно, большинство физико-химических и регуляторных сдвигов в процессе канцерогенеза так или иначе коррелирует с усилением ПОЛ во всех мембранных структурах клетки, неизбежно ведущим к нарушению многих связанных с ними ферментативных и неферментативных процессов.
Изменяются, в частности, содержание cAMP и соответственно интенсивности зависимых от него энергетических, синтетических и регуляторных процессов, которые при снижении уровня cAMP обычно способствуют пролиферации и, наоборот, тормозят её при возрастании концентрации cAMP. Эти закономерные прояв-ления обнаружены в основном в исследованиях 80-х годов XX века, и, как видно из рис. 16, падение уровня cAMP в клетке действительно приводит к различным преимущественно промитогенным эффектам. Что касается роли cAMP как ингибитора пролиферации большинства типов клеток, то она под-тверждена и в некоторых работах последних лет. В одной из них (Razavi et al., 1998) показано, что действие cAMP в ядре опосредуется факторами транскрип-ции, которые являются продуктами определённых генов. Таковым, в частности, считают индуцируемый cAMP ранний транскрипционный репрессор ICER-II. Последний блокирует клетки на границе фаз G2/M клеточного цикла. Он же ингибировал синтез ДНК в клетках опухоли гипофиза мыши и в клетках хо-риокарциномы человека, т. е. ICER-II признаётся опухолевым супрессором, который определяет антипролиферативную активность cAMP. Об экспрессии активного ICER-II при накоплении cAMP в Т-лимфоцитах человека и соот-ветственно подавлении пролиферации и эффекторных их функций было сооб-щено в другой работе (см. Bodor et al., 1996).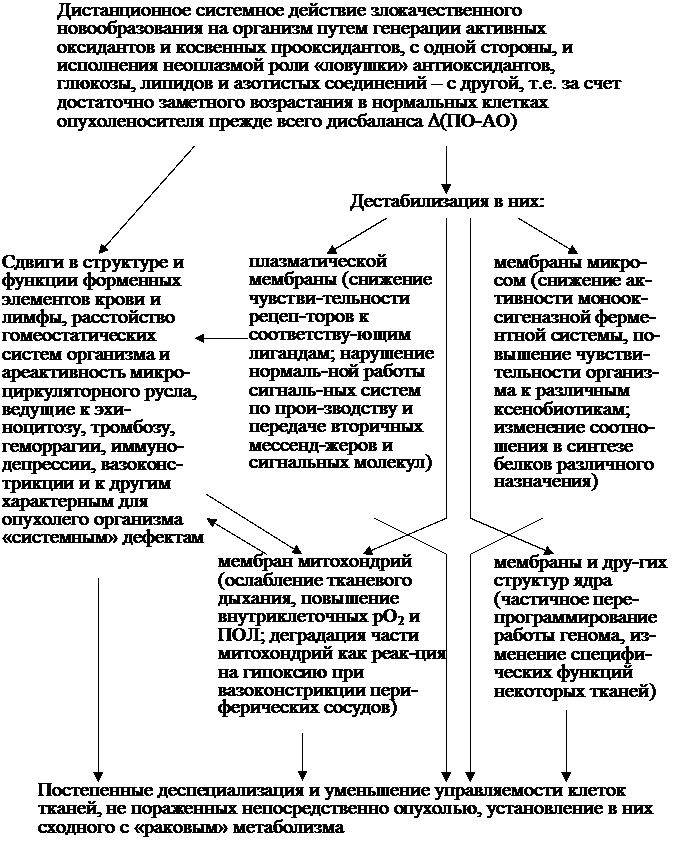 | |||
| |||
К развитию окислительного стресса при канцерогенезе причастен, несом-ненно, процесс ангиогенеза. В уже сформировавшемся очаге неоплазии ангиогенная активность поддерживает опухолевый рост «вширь», а в гиперплази-рованных участках тканей способствует перерождению этих пролифератов в новообразования. В числе идентифицированных ангиогенных факторов большинство является митогенами и лишь некоторые отвечают за формирование сосуда. Атакуя кровоснабжение неоплазм, можно в принципе получить лечебный эффект. В этой связи небезынтересно, что известные в настоящее время потенциальные ингибиторы ангиогенеза с различными механизмами действия проходят клинические испытания, и, по крайней мере, один из них находится под контролем гена супрессора опухоли р53 (Folkman, 1996).
Пролиферации и дисдифференцировке клетки способствует также одно-стороннее срабатывание в ней «позиционных» регуляторов. Таковыми представляются процессы «ассоциации – диссоциации» определённых субклеточных элементов (ферментов, рибосом, белков-репрессоров, гормон-рецепторных комплексов и др.) с мембранными структурами, матриксом или управляемым участком на молекулах биополимеров. На этот счёт в соответствующих параграфах главы приведены некоторые примеры.
Механизм многих изменений в ядерном геноме в норме и при канцерогенезе остаётся пока не ясным. Не исключено, что гены подсистемы управления клеточным циклом, в том числе определённые клеточные онкогены, имеют какую-то специфическую избирательную связь с ядерным матриксом по сравнению с остальными генами. Прикрепление их на период пролиферации к определённым участкам матрикса (как и некоторых других элементов клетки к соответствующим мембранам и скелетным структурам) целесообразно и необходимо со всех точек зрения для упорядоченной и надёжной реализации процессов транскрипции (прежде всего, на «митогенных» участках ДНК), а также синтеза и репликации последней. Возможно, это объясняет известный феномен амплификации протоонкогенов. Любопытно, что, согласно немногим сведениям (Eisenman et al., 1986; Эренпрейс, 1988), некоторые онкогены кодируют белки, связывающие определённые генетические элементы с ядерным матриксом, т. е. способствуют упорядоченной локализации этих элементов в пространстве ядра и последующим упорядоченным синтезам. Все другие участки генома, особенно мобильные генетические элементы, могут, вероятно, относи-тельно свободно отделяться от ядерного матрикса, спонтанно перемещаться и встраиваться в «чужие» области ДНК. Если предположить, что в связывании указанных участков с матриксом принимают участие также легкоокисляемые липидные компоненты, то при ПОЛ липидные связи могут нарушаться и тем самым создавать условия для транспозиции соответствующих фрагментов ДНК и спонтанных неточечных мутаций.
Кроме того, указанные перемещения генетических элементов, наряду с «неплановыми» переключениями в транскриптоно-триггерной системе клетки (вследствие, например, модуляции активности генома при перекисном окислении фосфолипидов, связанных с белками хроматина), могут быть ответственны за перепрограммирование генома опухолевой клетки и прогрессию неоплазм. Определённый вклад в эти нарушения вносят и непосредственные воздействия на ДНК АФК и продуктов ПОЛ. Большинство упомянутых изменений в ядре – события, на наш взгляд, вторичные по отношению к управляющим сигналам, поступающим из цитоплазмы как в норме, так и после воздействия на её структуры канцерогенных факторов. Последние в этом смысле негенотоксичны, а мутагенные свойства канцерогенов не имеют прямого отношения к механизму их канцерогенного действия (Lijinsky, 1989).
Другими словами, изменение нормальной клетки под влиянием кислородно-перекисного стресса является не просто ранним, но и необходимым этапом канцерогенеза. Последующее же повышение частоты мутаций и возникновение «мутаторного» фенотипа (Loeb, 1991) не есть достаточное, а в некоторых случаях даже необязательное событие канцерогенеза. В целом дискуссия об эпигенетических механизмах канцерогенеза, исключающих прямую модификацию или поражение яДНК соответствующими агентами и факторами, далеко не окончена. Об этом свидетельствуют и недавние обзоры работ по негенотоксиче-скому канцерогенезу. В последний могут вовлекаться различные потенциальные механизмы и клеточные процессы, в том числе ведущие к изменениям в регуляции роста клеток и экспрессии генов (Klaunig et al., 2000). Сообщается об идентификации нескольких новых ферментных комплексов, обратимо модифи-цирующих архитектуру хроматина путём метилирования, фосфорилирования и ацетилирования коровых белков нуклеосом. Некоторые из этих ферментов мо-дифицируют хроматин в норме, а в случае нарушения их функции ведут к искажению генной экспрессии и клеточной трансформации (Chakraborty et al., 2001). Становится всё более очевидным, что в канцерогенезе действуют 2 сос-тавляющие – эпигенетическая и генетическая (Лихтенштейн, Киселева, 2001).
С позиций развитых в данной главе представлений о механизме канце-рогенеза изменения в цитоскелете, плазматической мембране и внеклеточном матриксе достаточно логично увязываются с возникновением таких свойств злокачественности как инвазивный рост и метастазирование неоплазм. Проинвазивная и прометастазирующая роль окислительного стресса показана во многих исследованиях. Напротив, различные антиоксиданты часто проявляли себя как антиинвазивные и противометастазные средства. В числе таковых оказался, в частности, и cAMP, который, наряду с усилением митохондриального дыхания, синтеза ATP и ATP-зависимой полимеризации структур цитоскелета, стимулирует синтез молекул адгезии и белков щелевых контактов. В этой связи кажется естественным, что устойчиво повышенный уровень ПОЛ, дефицит ATP и cAMP в клетках неоплазмы создают, с одной стороны, условия для неконтролируемой их пролиферации, но, с другой, одновременно приводят к нарушению системы регуляции интегративных межклеточных взаимодействий и связей с внеклеточным матриксом. Немаловажно и более общее представление: врeменное снижение адгезивности и межклеточных контактов у стимулированных к размножению нормальных клеток, приуроченное к определённому этапу их митотического цикла, становится постоянным характерным признаком для злокачественных клеток.
Применительно к клеткам кроветворной ткани и некоторым типам форменных элементов крови тот же «кислородно-перекисный» подход представляется правомерным для анализа и обсуждения сущности различных видов лейкоза как одной из специфических форм онкологического заболевания. Наиболее реа-льными клетками-мишенями для лейкозной трансформации считаются полипотентные СКК и клетки-предшественники отдельных ростков кроветворения. Высокая чувствительность данной категории клеток к различным лейкозогенным факторам определяется, по нашему мнению, умеренно гипероксическими в них условиями, которые создаются, главным образом, за счёт малого числа действующих митохондрий. В норме такие условия необходимы, очевидно, для устойчивого протекания окислительного митогенеза в кроветворной клетке и обеспечения практически неограниченной способности её к пролиферации. Содержание митохондрий может быть низким как изначально, так и вследствие клеточного старения и соответствующего повреждения этих органелл. В любом случае такие клетки потенциально опасны в лейкозогенном отношении.
При лейкозной трансформации даже имеющиеся немногие митохондрии подвергаются дезорганизации, и по данному признаку лейкозные клетки не отличаются от неопластических другого вида. По названным выше причинам СКК и их потомки высокочувствительны в негативным сдвигам в дыхательном аппарате: в них «пролиферативный» дисбаланс ∆П (ПО – АО) сравнительно легко увеличивается до «лейкозогенного» ∆Л (ПО – АО) – аналога «канцерогенезного» дисбаланса ∆К (ПО – АО) для негемопоэтических клеток опухоли. Кислородно-перекисная концепция лейкозогенеза объясняет многие факты про-текторного действия различных антиоксидантов, среди которых в последнее время часто упоминаются, например, ресвератрол и некоторые другие флавоноиды (см. п. 7.1.2).
Представление об умеренном и повышенном уровнях ПОЛ соответственно в пролиферирующих нормальных и опухолевых клетках вместе с другими положениями кислородно-перекисной концепции онкогенеза позволяют понять, казалось бы, парадоксальный феномен. Имеются в виду те факты, когда одни и те же прооксидантные воздействия, создающие гипероксию и (или) повышающие уровни АФК и ПОЛ мембран, по отношению к нормальным пролиферирующим клеткам часто ведут себя как онкотрансформирующие факторы, а для опухолевых клеток они выступают как цитотоксические. Примерами воздействий, приводящих нормальные и опухолевые клетки к указанным двум разным исходам за счёт того, что в них реализуются соответственно переходы ∆П (ПО – АО) → ∆К (ПО – АО) и ∆К (ПО – АО) → ∆А2 (ПО – АО) или ∆К (ПО – АО) → ∆Ц (ПО – АО), могут служить гипероксия, радиация, гипертермия, перекисные соединения, ингибирование эндогенных антиоксидантов и клеточного дыхания, действие некоторых антибиотиков и др. Что касается применения в лечебных целях антиоксидантов, то отдельные из них (прежде всего, антира-дикального действия) также обладают двойственным эффектом. При терапии неоплазм такими антиоксидантами возможна инверсия эффекта с протектор-ного в стадии инициации на опухолепромоцирующий и (или) генотоксический для уже инициированных клеток (Hartman, Shankel, 1990).
Противоположное действие антиоксидантов на разных стадиях канцерогенеза может быть опять-таки связано с различием в них уровней пероксиге-нации. На стадии инициации, когда интенсификация свободнорадикального окисления только начинается и антиокислительная активность в клетках ещё низка, эндогенные антиоксиданты препятствуют возрастанию в них дисбаланса ∆ (ПО – АО) до уровня канцерогенных значений ∆К (ПО – АО). Однако на стадии максимального «разгона» пероксигеназных процессов в активно растущих опухолевых клетках введение антиоксидантов, несмотря на повышенную к этому времени антиокислительную активность, может привести к обратному эффекту. Такой исход возможен ввиду реального в названных условиях окисления самого ингибитора и образования при этом новых активных свободных радикалов, участвующих в реакции продолжения цепи. Результат здесь будет зависеть также от дозы вводимого антиоксиданта: небольшие дозы (ниже некоторого критического значения) расходуются в основном путём самоокисления, усиливая процессы пероксигенации, большие же – способны снизить или даже потушить цепные реакции окисления в опухолевых клетках.
Примером названной двойственности эффекта могут служить данные о дей-ствии на рост клеток плоскоклеточного рака полости рта человека витамина Е: физиологические концентрации его (0,001-50 мкМ) стимулируют, а фармакологические (100-154 мкМ) – ингибируют рост тех клеток (Elattar, Virji, 1999). В первом случае токоферол частично, по-видимому, самоокисляясь, поддерживает «канцерогенезный» дисбаланс ∆К (ПО – АО) и одновременно не допускает повышение последнего до проапоптозного диапазона ∆А2 (ПО – АО). Во втором же случае витамин Е снижает ∆К (ПО – АО) до уровня, приводящего, скорее всего, к апоптозу А1, а иногда и к нормализации метаболизма. Указанные выше представления оказались полезными для объяснения механизма противоопухолевого действия и систематизации значительного числа лечебных мероприятий (см. главу 7; Лю, Шайхутдинов, 1991).
Таким образом, в аспекте эволюционного учения и первопричинности мно-гие дегенеративные изменения и неинфекционные болезни, включая процесс старения и рак, могут являться только следствием аэробного существования живых организмов (Лю, Ефимов, 1976; Harman, 1984; Kang et al., 1994; Лю, 2001). Последние, адаптируясь к этим токсическим условиям, «внедрили», прежде всего, на клеточном уровне многоступенчатую антиоксидантную систему, в которой наиболее значимую роль играют митохондрии. Однако уязвимость и чувствительность их к различным повреждающим факторам резко снижает эффективность всей антиоксидантной системы клетки, что является основной первопричиной нарастания в ней гипероксии и оксигеназного стресса со всеми вытекающими отсюда негативными последствиями. В этих условиях усиление тем или иным способом различных ступеней антиоксидантной защиты вполне оправдано в качестве геропротекторных, противовоспалительных и антиканцерогенезных мероприятий, как наиболее адекватных состоянию избыточной пероксигенации. К сожалению, при развившихся уже опухолях антиоксидантная терапия в большинстве случаев не приводит к полной их регрессии. Поэтому решающее, на наш взгляд, значение приобретают оксидеструктивные методы лечения неоплазм (см. п. 7.2), к которым активно растущие участки последних «подготовлены» ввиду присущих им биоэнергетических и других особенностей, обсуждавшихся в данной главе монографии.
Еще по теме клеточная пролиферация:
- Молекулярный шаперон HSP70 и скорость клеточной пролиферации в онкогенезе и риске развития метастазов при инвазивном раке молочной железы
- Роль маркеров пролиферации
- Фаза восстановления или пролиферации
- Пролиферация клеток и формирование мембран
- Тема № 7. Факторы апоптоза и пролиферации
- 3.6. Биоэнергетические аспекты механизма пролиферациии дифференцировки клеток
- 5.2.2. Препараты, преимущественно воздействующие на пролиферацию и дифференцировку В-лимфоцитов.
- Особенности пролиферации клеток из зон НПА и ВПА (исследование с помощью кривых роста)
- Особенности пролиферации клеток из зон НПА и ВПА (исследование с помощью радиоавтографии)
- Оценка динамики зрительных функций при отслойках сетчатки и на ранних стадиях витреоретинальной пролиферации
- Тема № 6. Белки - маркеры клеточного цикла
- Ингибиторы синтеза белка и клеточного деления