КЛЕТОЧНЫЙ И ТКАНЕВОЙ УРОВНИ АДАПТАЦИЙ У ГОМОЙОТЕРМНИХ ОРГАНИЗМОВ
Поскольку для гомойотермных организмов характерно поддержание постоянства внутренней среды, явления клеточных и тканевых адаптаций, казалось бы, должны занимать незначительное место в общей картине приспособления к различным факторам среды и условиям существования.
Однако (как это будет показано в последующем изложении) представление о гомеостазисе как о строгом постоянстве физических и химических условий во внутренней среде организма (крови, тканях, отдельных системах органов) может служить лишь схемой, практически никогда не выявляющейся в условиях природных отношений организма. Это положение в равной мере относится к температуре отдельных частей тела, pH тканей, содержанию воды в клеточных элементах и в межклеточных пространствах, напряжению кислорода в тканевых капиллярах и т. д., это касается также и поддержания постоянства химического состава организма — процесса питания, который далеко неодинаково обеспечивает расход вещества в отдельных органах и системах.Так, голодание неодинаково сказывается на весе отдельных органов н систем (Пашутин, 1902).
В последнее время изучалась возможность изменения тканевого дыхания у разных животных при голодании. Оказалось, что полное голодание вызывает не только значительное падение веса отдельных тканей, но и ограничение жизнедеятельности тканей, что находит свое
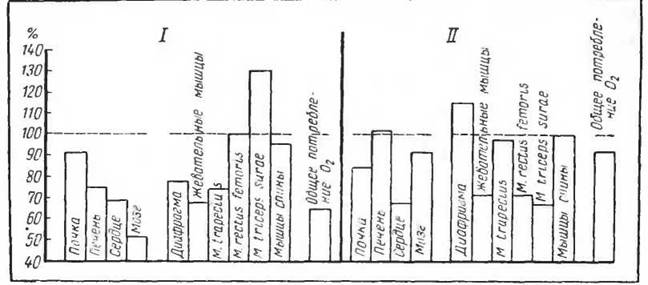
Рис. 3. Изменения тканевого дыхания при голодании (по Слони- му, 1968). / — краснощекий суслик; II— белая крыса
отражение в снижении их энергетического расхода. На рисунке 3 приведены некоторые данные, полученные на голодающих крысах и краснощеких сусликах.
Неоднородная реакция дыхания тканей при голодании указывает па особую тканевую специфичность и на существование специальной регуляции обмена веществ при голодании и при питании.
В пользу этого говорит и тот факт, что наличие жирового депо в значительной мере сглаживает изменения тканевого дыхания при голодании.В истории изучения клеточных адаптаций значительную роль сыграл постулат Пфлюгера (Pfliiger, 1878), установившего, что количество энергии, вырабатываемой клетками тканей тела организма, не зависит от доставки к ним кислорода или пищевых веществ. Это положение Пфлюгера оказало большое влияние на последующие работы и развирало мысль о том, что потребление кисло* 38
рода клеточными изолированными элементами, а также изолированными тканями не зависит от общего уровня обмена животного. Ткани пойкилотермных и гомойотермних животных (лягушка, кролик, мышь, собака) при исследовании in vitro потребляют при одинаковых температурах равное количество кислорода на 1 г веса ткани (Grate, 1928). Отсюда был сделан весьма важный вывод, что уровень общего обмена веществ определяется не свойствами самих клеток и тканей, а сложным нервно-гуморальным аппаратом регуляции их деятельности. Последующие исследования ввели значительные поправки в эти широко известные данные Графе. Изолированная мышечная ткань крыс и мышей обладает при одинаковых температурных условиях более высоким уровнем обмена веществ, нежели ткань более крупных грызунов — кроликов (Krebs, 1950; Bertalanffy a. Estvick, 1953). В этих же исследованиях обнаружилось, что в пределах одного вида изолированная мышечная ткань животных разного веса потребляет одинаковое количество кислорода. Следовательно, положения Графе оказываются справедливыми для животных разного веса в пределах одного вида, но не подтверждаются для животных разных видов с разной относительной поверхностью тела. Берталаифи и Пироцинскнй (Bertalanffy a. Pyrozynski, 1953) указывают, что внутривидовые различия в уровне основного обмена, связанные с относительной поверхностью тела, обусловливаются влиянием нервной и гуморальной систем. У разных же видов можно обнаружить заметные и притом стойкие различия тканевого дыхания, зависящие от внутриклеточных особенностей тканей.
Ряд исследований установил неодинаковую реакцию тканей тела на охлаждение и нагревание. У белых мышей наблюдалось значительное повышение потребления кислорода тканью печени после кратковременного (30- минутного) пребывания животных при низкой температуре ( + 5°С), у шестинедельных лабораторных крыс наблюдается после 10 и 26-дневного пребывания при температуре 4-5° С повышение потребления кислорода всеми тканями тела, за исключением ткани головного мозга (Weiss, 1954). Когда таких же крыс содержали 5 дней при температуре +12° С, повышения потребления кислорода тканями не наблюдалось. Таким образом, несмотря на последующую изоляцию тканей от организма,
можно наблюдать повышение газообмена в тканях после воздействия холода.
В условиях значительной гипотермии наблюдается очень большое снижение (до 22% ОТ исходного уровня) потребления кислорода тканями мозга (Калихман, 1953). В то же время снижение газообмена в ткани печени было значительно меньшим даже при температуре
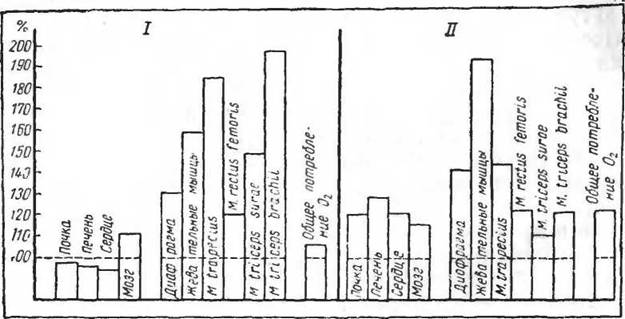
Рис. 4. Изменения тканевого дыхания при адаптации к холоду (по Исаакян, Макаровой, Щегловой, 1969). I— индийская крыса
(Nesokia indica); II — белая крыса
+ 15°С. Следовательно, при предварительном охлаждении целого животного можно наблюдать значительные сдвиги общих энергетических затрат тканей. Каждая ткань имеет, по-видимому, свои параметры в этих изменениях обмена веществ. Интересно отметить, что газообмен ткани мозга при охлаждении до температуры тела +27° С не повышается, тогда как в тканях печени и мышц это повышение очень значительно (Калихман).
Длительная адаптация (6 недель) к холоду приводит к значительному повышению тканевого дыхания у хорошо адаптирующейся белой крысы (во внутренних органах). У тропического вида индийской крысы {Nesokia indica) повышение тканевого обмена возникает и поддерживается только в скелетной мускулатуре. Эти сдвиги не обеспечивают у тропических грызунов поддержания постоянной температуры тела при охлаждении (рис.
4).Изучение реакций организма на тканевом и клеточном уровнях открывает широкие перспективы для понимания стойких изменений в организме, возникающих под влиянием факторов внешней среды. Это хорошо иллюстрируется изменениями клеточных элементов скелетной мускулатуры при воздействии холода.
Общеизвестно, что скелетная мышца по своей микроструктуре не однородное образование; отдельные ее волокна обнаруживают различную способность к длительным тоническим сокращениям, чувствительность к ацетилхолину, отличаются по содержанию миоглобина и т. д.
Оказалось, что сильно реагирующие при охлаждении организма мышцы — трапецевидная и мышца бедра у крыс (см. стр. 184) —не являются однородными по толщине мышечных волокон, снабжению этих волокон капиллярами и по содержанию окислительных ферментов (сукциндегидрогеназа). По содержанию окислительных ферментов мышцы состоят как бы из трех клеточных популяций. Степень васкуляризации волокон, содержащих большое количество фермента, в полтора раза выше, чем волокон, в которых фермента мало. Было выявлено и распределение волокон с различным содержанием окислительного фермента. В мышцах крысы обнаружено своеобразное, расположенное в центре мышцы ядро, интенсивно окрашенное и содержащее много фермента, и периферическая часть с малым содержанием фермента. Такое распределение волокон с разным содержанием фермента не является, однако, постоянным. Экспозиция крыс в течение 60—100 дней при температуре + 5° С привела к некоторому изменению состава клеточных популяций, количество интенсивно окрашенных волокон возросло (Дерибас, Филипченко, Шошен- ко, 1966).
Механизм тканевых адаптаций физиологических процессов можно проследить не только по изменениям общей жизнедеятельности клеточных элементов, общим выражением которой является потребление кислорода. Немаловажное значение приобретают в этом случае и отдельные стороны обмена веществ, и связанные с ним накопления отдельных потенциалов клеточных энергетических превращений.
Так, например, под влиянием холода как в эксперименте на крысах (Page et Babineau, 1954; Pavlovic, 1954), так и у животных — обитателей полярных областей (Шварц, 1959; Ливчак, 1960) возрастает содержание гликогена в печени. Под влиянием охлаждения возрастает количество аскорбиновой кислоты у крыс и обезьян (Dugal et Therien, 1947; Dugal et Fortier, 1952). У обского лемминга и большой узкочерепной полевки (полярные виды) по сравнению со степной пеструшкой (степной вид) наблюдалось повышенное содержание аскорбиновой кислоты в тканях почки, печени (Ливчак, 1960). В этой же работе установлены прямые взаимоотношения между потреблением кислорода на 1 г ткани целого животного и содержанием аскорбиновой кислоты в ткани почки. Эти данные хорошо иллюстрируются следующей таблицей.Таблица I
Интенсивность потребления кислорода и содержание аскорбиновой кислоты в почках у грызунов (по Ливчак, 1960)
| Вт жив л ног» | П )тр?бл?ние гри 163 С (лг'ч) | K ицон гашія А К в почках (мг%) |
| Обский лемминг (Lemmus obensis) | 3.6 | 11.2 |
| Узкочерепная полевка (Microtus | ||
| gregalis)....................... .............................. | 4.9 | 13.4 |
| Степная пеструшка (Lagurus lagu- | ||
| rus)............................................................. | 7,о | 20.1 |
В условиях гомойотермии значительные колебания температуры тканей отсутствуют, однако в ряде случаев приходится считаться с непосредственным влиянием холода на тканевые процессы.
Понижение температуры конечностей имеет место у большинства полярных млекопитающих и птиц.
Это находит свое отражение в интересных данных об особенностях точки плавления жира в разных частях тела полярных животных, (Irving, Schmidt-Nielsen a. Abraham- sen, 1957). Оказалось, что жир тканей внутренних органов имеет точку плавления +38—50° С, в суставных сумках стопы и пальцев — ниже +10° С (у лисицы, собаки и северного оленя около 0—5°С), что совпадает с данными для зимоспящих животных. У тропических животных точка плавлення жира конечностей много выше.Имеются довольно многочисленные данные, характеризующие термоустойчивость живых тканей у пойкило- термных организмов (Ушаков, 1955, 1956). Обнаружены химические особенности белков мышечного волокна. Это доказывается различной устойчивостью мышц к нагреванию при действии аденозинтрифосфатазы. К сожалению, данных для млекопитающих и особенно для мышц в целом организме, функционирующих при различных температурах (например, для мышц конечностей или мышц туловища), пока пет.
Для свойств пищеварительных ферментов [1] были установлены как факты изменения их видовых особенностей (например, у плотоядных и растительноядных организмов), так и при длительном питании всеядных животных растительной или животной пищей (Уголев, 1958а). Это оказалось ярко выражено в особенностях ферментов, расщепляющих растительный и животные углеводы (ферментативной активности по крахмалу и по гликогену).
Весьма значительные изменения в химических процессах в тканях наблюдаются в связи с особенностями снабжения их кислородом. С этой стороны особый интерес представляют данные о содержании миоглобина, указывающие на то, что в мышцах млекопитающих, устойчивых к функциональной гипоксии (горные животные, ныряющие животные, животные, выполняющие интенсивную мышечную работу) количество миоглобина повышено (Hurtado, Rotta, Alerino, Pons, 1937; Irving, 1939; Robinson, 1939; Clark, Criscuolo, Coulson, 1952; Верболо- вич и сотр., 1955, 1956).
Многлобин во всех случаях является веществом, способным связать дополнительное количество кислорода в самой мышечной клетке, активировать окислительные процессы с использованием кислорода в момент ограничения доставки его извне (например, при нырянии) или при интенсивной мышечной работе, когда снабжение кислородом через дыхательно-циркуляторную систему оказывается меньшим, чем кислородный запрос работающих мышц.
Однако тканевые клеточные процессы в условиях ограничения доставки кислорода изменяются не только в связи с увеличением содержания миоглобина как вещества, создающего известное депонирование кислорода. Как уже указывалось выше, постулат Пфлюгера предусматривает независимость интенсивности тканевого дыхания от содержания кислорода в окружающей среде. Следовательно, процесс приспособления к недостатку кислорода должен приводить с этой точки зрения к более полному поддержанию уровня окислительного метаболизма независимо от условий снабжения кислородом.
Потребление кислорода кусочками тканей сердца, печени, головного мозга, диафрагмы, взятыми от акклиматизированных мышей (поднимавшихся систематически в барокамере), немного повышается в среде атмосферного воздуха и резко возрастает при снижении давления кислорода до 1,5—1,7% (Барбашова, 1960). В последнем случае потребление кислорода тканью сердца составляет 202% для мышей и 218% для крыс, почечной тканью соответственно 190 и 191%, тканью головного мозга — 212 и 219% и диафрагмой у крыс—185%- Следовательно, у акклиматизированных лабораторных крыс и мышей возрастает способность тканей поддерживать высокий уровень потребления кислорода в бедной кислородом среде. Такой высокий уровень потребления кислорода имеет место только при крайнем ограничении его доставки в ткани. Близкие данные были получены в работе Сэндстрема и Михаэлса (Sundstroem a. Michaels, 1942) и некоторых других авторов, хотя, по-видимому, для различных тканей условия, в которых поддерживается относительно повышенный при недостатке кислорода окислительный метаболизм, могут быть различны.
Имеются данные, свидетельствующие об изменениях в состоянии тканевых окислительно-восстановительных систем (цитохрома, цитохромоксндазы). Содержание цитохрома, по данным той же работы 3. Барбашовой, в мышцах далеко не всегда возрастает в процессе акклиматизации к высоте. Содержание сукциноксидазы у акклиматизированных кошек, крыс и мышей, однако, возрастает по сравнению с контрольными животными. Особенно резко возрастает активность этого фермента в условиях пониженного содержания кислорода в среде, окружающей мышечную ткань. При достаточном снабжении ткани кислородом активность сукциноксидазы у акклиматизированных и неакклиматпзированных животных не отличается (Крепе, Вержбннская, Ченыкаева, Чирковская, Гавурина, 1956). Таким образом, между характером потребления кислорода в бедной кислородом среде и активностью сукциноксидазы существует прямой параллелизм, характеризующий состояние тканей животных, акклиматизированных и неакклиматизированных к пониженному барометрическому давлению.
Кроме изменений цитохромной системы под влиянием высотной акклиматизации, в тканях обнаружены сдвиги ряда других ферментных систем. Так, обнаружено увеличение желтого фермента и дегидразы (Delachaux et Tissieres, 1946), повышение активности сукциндегидроге- на н трифосфатазы (Tappan, Reynafarje, van Potter a. Hurtado, 1957). Резко возрастает содержание в тканях карбоангидразы (Вержбинская, 1946; Стрельцов и Хазен, 1946; Пигарева, 1947 и др.). Фермент карбоангидраза находится в тесной связи с интенсивностью обмена веществ. Значение его заключается в ускорении удаления СОг из тканей. Это особенно ярко выражено в хрусталике глаза, где кровеносные сосуды отсутствуют, и в сетчатке глаза, где васкуляризация выражена слабо, несмотря на интенсивный обмен веществ (Четвериков, 1948; Kauth a. Sommer, 1953).
Под влиянием повторной и длительной гипоксии может изменяться также соотношение между анаэробной и аэробной фазами метаболического процесса в тканях, особенно в мышечной ткани. Повышение анаэробного гликолиза в эмбриональных тканях развивающегося в условиях гипоксии куриного яйца наблюдала Т. Ф. Комарова (1952). Такое же явление было обнаружено в мышцах акклиматизированных к высоте животных (Бар- башова, 1960), а также после тренировки к скоростным, сопровождающимся острой кислородной недостаточностью мышечным напряжениям (Яковлев, 1950, 1955).
Повторное воздействие гипоксии, например при ане- мизации конечности, приводит к изменениям местной тканевой возбудимости (хронаксии), указывающим на наличие тканевых процессов адаптации к гипоксии. Резкое падение возбудимости становится значительно менее выражепым после 15-минутной анемнзации конечности (руки человека), производившейся ежедневно в течение месяца. В этом случае имеет место очень сложная картина адаптации как рецепторов тканей, так и, возможно, самих мышечных элементов.
Однако едва ли приведенные выше еще отрывочные данные в настоящее время могут исчерпать всю сложную картину тканевых адаптаций млекопитающих, возникающую под влиянием факторов внешней среды. Устойчивость целого организма к гипоксии, наблюдающаяся в результате высотной акклиматизации, устойчивость тканей животного к низким температурам, свойственная многим арктическим животным и птицам, далеко выходят за пределы значения описанных выше частных физико-химических механизмов. Следовательно, различные стороны явлений такой устойчивости, хотя и однородные по их внешним проявлениям, не могут быть объяснены известными физиологическими механизмами.
Адаптации к изменению содержания воды и осмотического давления складываются из процессов удержания внутриклеточной и межклеточной воды. Содержание воды в тканях большой песчанки представляется в следующем виде (см. табл. 2).
Таблица 2
Содержание воды в органах большой песчанки при питании растительным кормом разной влажности (по Арешевой и Щегловой, 1961)
| Влажность естественных кормов | Вес тела (г) | Сердце | Печень | Почка | Мышцы | Кожа |
| 73—76% | 90,3 | 81,4 | 74,2 | 79,9 | 78,1 | 64,5 |
| 43-47% | 93,5 | 79,2 | 73,2 | 79,0 | 77,7 | 67,1 |
При уменьшении влажности корма с 73—76% ДО 43— 47% содержание воды в тканях ряда органов — печени, почке, сердце, скелетной мускулатуре — уменьшается. В то же время в коже содержание воды несколько возрастает. Это указывает на несомненную роль последней в качестве водного депо организма.
Содержание воды в мышцах овец и верблюдов в условиях пустыни Каракумы претерпевает значительные сезонные изменения (Кузнецов, 1947). Так, содержание воды в феврале равнялось в среднем 76,65%, в мае — 77,65% (до 89,05%), в июне — 77,82%, в ноябре — 76,28%- Увеличение влажности корма в весенние месяцы сопровождается и возрастанием содержания воды в скелетных мышцах. Следовательно, скелетная мускулатура, составляющая значительную часть тела (по весу), является у этих животных водным депо. Его значение у разных млекопитающих в процессе приспособления к условиям недостатка воды еще неясно. Очень важен вопрос об образовании в процессе обмена веществ так называемой метаболической воды.
У сусликов обнаружено повышенное образование метаболической воды в процессе окислительного метаболизма (Ковальский, Шульгина и Ржешевский, 1951). По их данным, этот процесс может быть искусственно усилен в условиях ограниченной доставки воды в организм.
Итак, тканевые процессы гомойотермных организмов подвержены глубокой адаптации, направленной подчас на изменение ряда физико-химических клеточных механизмов, но обеспечивающей существование животного в специальных, а иногда трудных для жизни условиях среды.
Тканевые (клеточные) адаптации связаны не только с прямым воздействием факторов внешней среды, но и отражают особенности регуляции функций отдельных систем целого организма. Так, например, ткани зимоспящих животных нечувствительны к глубокому охлаждению только при виключений терморегуляции, т. е. в состоянии зимней спячки. Однако у бодрствующего животного сильное охлаждение вызывает такие же картины нарушения функций, как и у других млекопитающих.
Еще по теме КЛЕТОЧНЫЙ И ТКАНЕВОЙ УРОВНИ АДАПТАЦИЙ У ГОМОЙОТЕРМНИХ ОРГАНИЗМОВ:
- №3 Организм человека как единая биологическая система. Уровни организации человеческого организма: молекулярный, клеточный, тканевой, органный, системный, организменный.
- Организм как тканевой симбиоз
- 7.Уровни адаптации
- 1. Представления о процессе адаптации организма.
- Уровни регуляции клеточного ответа.
- Уровни организации человеческого организма
- Современные методы и средства для оценки адаптационных возможностей и уровня здоровья организма человека
- 14.3 Характеристика основных психических состояний человека по уровню активации организма
- Тканевая (гистотоксическая) гипоксия
- Зависимость уровня экскреции белка ТНР от уровня потребляемой соли, риск ухудшения натрийуреза.
- Костно-тканевая проводимость.
- Исследование уровня притязаний и уровня самооценки у детей по Дембо-Рубинштейн в модификации А.М. Прихожан («Лесенка»), оценка результатов.
- Подавление тканевой несовместимости при кератопластике
- 27. Профессиональная адаптация. Причины трудностей профессиональной адаптации
- Паренхимноклеточная и тканевая эмболия
- Тканевой материал.
- Разобщители тканевого дыхания
- Тканевой обмен слюнных желез рта