3.3 Действие Rpf на клеточную стенку Л/, tuteus
При изучении воздействия Rpf на культуру М. luteus наблюдается одно любопытное явление. При выращивании на бедных средах, таких как лактатная, бактерии микрококка склонны образовывать крупные агрегаты, что, скорее всего, является ответом на недостаток питательных веществ.
Нами было отмечено, что при добавлении Rpf к культуре, пребывающей в подобном сильно агрегированном состоянии, происходила частичная дезагрегация клеток и размеры агрегатов в целом уменьшались. Так> если до воздействия белка агрегаты могли насчитывать сотни клеток, то после добавления Rpf основная часть агрегатов состояла уже максимум из двух трех десятков клеток, и количество единичных бактерий возрастало.Предположительно, данное явление может быть связано с некой гидролитической активностью, способствующей дезагрегации клеток за счет модификации (разрушйдоя) неких связей в поверхностных структурах микрококка (клеточной стенке). Наличие «ферментов дезагрегации» действительно было обнаружено для ряда бактерий. Для того чтобы проверить наши предположения, мы решили выяснить оказывает ли Rpf какое-либо воздействие на клеточные стенки микрококка. Для этого измеряли во времени оптическую плотность препарата лиофилизированных клеток микрококка после добавления к ним рекомбинантного Rpf (рис. 17).
65
20 ; ю I
5 5 15 25 35 45
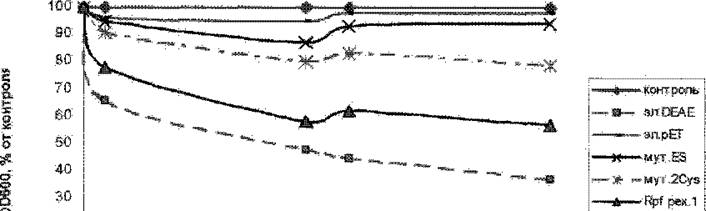
Pucl IT Изменение оптической плотносш препарата л шфи шифрованных клеток М &&ш в 20мМ трие-буфере рН 8 иод дейсшем препарата решмбинантного Rpf, а так же рекомбинантных Kpf белков, подвергнутых саш"-наггравленному мугагхншу. Эл,ОКАН -злвдат, содержит дативный Rp^ выделенньай на колонке DHAE щ оупернатанта. Эл рЬТ -контрольное выделение ржомбинантного Rpf из штампа К coiL содержащего пустой вектор без гена Rpf Myx.ES ™ решмбинаншый Rpf с "тмеиой шутами на -33 в RS-фрагментс на глутаминовую к-ту, MyxCys - Rpf с заменой обоих цисте! шов в тжерватшшш ломенс (Сук 50 на Lys, Cys 110 на Thr).
Rpf р«Л - препарат рФшда&гшнтного Rpf Концентрации ретомбшантного Rpf - 15 мкг/мл, ДИК» конструкции для экспрессия Rpf с замененными! аминокислотами были любезно предоставлены ПМукатиоловой. Bee стадии очистки зиутантных форм Rpf шответствивали стандартному иротоголу полу^шш нешмшбниой формы Rpf. Из рнс< 17 ш*диоч *гго при добаадешш реюмбииаятного Rpf к ярегарату лшфилшйро&атш клеток микрококка происходит падение оптической плотности, в конечном итоге нритодятее к уменьшению оптической плотности примерно до 60% от начальной. Можно предположат^ что66
данный эффект является отражением гидролитической активности, присущей Rpf. Столь медленное падение оптической плотности, скорее всего, связано со специфичностью Rpf Вероятно, он гидролизует какие-то связи,' расщепление которых не приводит к глобальным изменениям в структуре крупных частиц клеточной стенки, ведущим к их полному распаду, что могло бы привести к существенному и быстрому падению оптической плотности, как это происходит под действием лизоцима, Скорее всего, действие Rpf, в отличие от лизоцима, не приводит к к тотальному гидпролизу клеточной стенки, а модифицирует структуру составляющего'., ее пептидогликана, разрушая некоторые связи и делая ее более рыхлой. Возможно, именно из-за подобного воздействия на клеточную стенку белок обладает способностью дезагрегировать клетки микрококка- явление, которое было отмечено выше.
При компьютерном анализе последовательности консервативного домена Rpf специальные программы обнаруживают сходство третичной структуры с так называемым лизоцимо-подобным фолдом, образуемым характерно расположенными в пространстве тремя альфа спиралями. (Рис, 18). Действиетельно, при моделировании возможной структуры консервативного домена наиболее веростной структурой оказалась таковая .близкая к структуре фрагмента лизоцима в районе о1 активного центр (данные любезно предоставлены д-ром А.Мурзиным- Великобритания) (Рис.18).
67
 |
| ..:■*■ * |
| ..■■:v ." ":3C 'JSft-Y- Jijfcugttj ■ . :-.v »:f-y:. '.vkH ,здь*ё |
fm*, Ш. Компьютер*:*.!^ модсж арукчур куршюго лшшкша (слева) и консерваишюш ломена Щй (иредСкглзшдкО (oupmni). D нижней >шетп рн'суик:). изображающего лиюцнм хорошо видны грн и-аш;пьли: яидпонп'^ся няр;нсгерш>?м&& док лтого (юлка. Пграша - па рнеутже предетниен кепссрштиЕДп.ш домен RpL где так же кидшл грп а-спир;;^и, ичлпмнсс распою же иве кеп-орых похоже на такпг;],1- imoixvuti.
\гл неак \\)срл:гпп№. участаунчпдх в мтжолише гслетолноп етенкгг, щпи'ше лшощшо-нодоегшх с груктур характерно дги! некоторых лшн'геекпх трапепшкозш:;п„ фермелтоп, моделирующих л& снег гидролиза клеточную стопку млгерооргапюхюв (Рис. 21) UIoUjc. 1995, 'I'himnL'^n сА n\; !99\). '-V;-^ ("пики ^о:яютс& пя^терма'^.гымй мура^идачпми, Чг№Йстштгшшл\ш н ме_1^бо1:и1ме б^^герпалыюй хлошчпоп егенгш во время рогд! и зепения> а гак же в процессах cvr//v,uum\ с :Hipyoei?iiiocr;:*o. ^сак:1ШГк кшаличаруе^ая аднми (Ямками, еосчппт и разречангш И-1Л глико'зпдпых ^з^язей пептпдоглш^'ла клеточной стсг.ки, еппр я",1.тшюм с сбраюгшшеа? 1,6» ангидрому^олегпидс^ ((Th'.inmsu. •М al, 1W1, ПдЕыи* н aL, ИНН*. Лцзошшы, прип;5дпежгшш€ к *ему тс супсрсс^:ейС1^) гтакашя пгфоткод да образуют ih речултлтне свое а реакции 1.6-аш^идромужгилпилок и обладают м^псс снециЛн uvaw чспстссм, лто делас1! их дейегше на
68
клеточную стенку более разрушительным (Engel et al., 1991). Под действием литических
трапсгликсзилаз бактериальная клеточная стенка не разрушается полностью (как в случае с лизоцимом), а становится более рыхлой и проницаемой. Благодаря этому, многие бактериофаги (фаг Т4, фаг Т7 и другие) используют именно эти ферменты для проникновения в клетку хозяина, поскольку после инфекции клетка не должна терять жизнеспособность (Lehnherr et al.51998).
В связи со всем вышеизложенным мы предполагаем, что Rpf является ферментом, участвующим в гидролитической модификации клеточной стенки.
Существует несколько типов ферментов, для которых характерна подобная активность (рисЛ9): лизоцимы (мурамидазы), эндопептидазы, амидазы, карбоксипептидазы и литнческне трансгликозилазы. Пока мы не можем однозначно определить, к какому типу ферментов можно отнести наш белок. Представляется, что Rpf вряд ли можно отнести к лизоцимам несмотря на его третичную структуру, так как М. tuteus— микроорганизм чрезвычайно чувствительный к действию лизоцима и малейшее его добавление в среду приводит к полному лизису клеток. У нас не существует каких-либо указаний на то, что Rpi является амидазой, анализ его последовательности не выявляет его сходства с этим типом ферментов.69
m-Aapm-Wfe I
GIcNAc -MurNAc -GlcNAc -MurNAc -GlcNAc -MurNAc-anh
I -*— 3 3* —■»- ' Transpeptldase
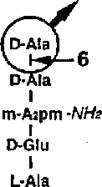 L-Ala L-Ala
L-Ala L-Ala
I л I
| D-GIu Г |
D-GIu 1
m-Aspm -Л/Н»
m-Aapm - D-Ala i r
^ l I
| i . |
D-Ala m-Aspm-WHs °"А,а
I
L-A!a
>£*i~
- GlcNAc -MurNAc -GlcNAc -MurNAc^# 41-» GlcNAc -MurNAc -•
L-Ala ^-^V
1 D-GIu
I m-Aapm-NH?
I D-Ala
Рис. 19. Ферменты клеточной стенки и их действие на муреин у Е. coli (по Holtje, 1995). GlcNAc -N-ацетилглюкозамин, MurNAc - N-ацетшшурамовая кислота, MurNAc-anh (I—>6)ангидромурамовая кислота. Связи, гидролизуемые соответствующим ферментом; 1 - N-ацетилглюкозаминидазы, 2 - литические трансгликозилазы, 3 - К-ацетилмурамил-Ь-аланин амидаза, 3* - К-ацетилдегидромурамил-Ь-аланин амидаза, 4 - DD эндопептидаза, 5 - LD карбоксипептидаза, б - DD карбоксипептидаза.
В пользу того, что он является литической трансгликозилазой, указывает наличие гомологии в третичной структуре консервативного домена Rpf и некоторых литических трансгликознлаз.
Так^же для некоторых трансгликознлаз характерен LysM домен, служащий для прикрепления к клеточной стенке, который присутствует и у Rpf. Как уже отмечалось выше, литические трансгликозилазы не разрушают клеточную стенку полностью, а лишь делают ее более рыхлой и проницаемой. Вследствие этого кинетика разрушения препаратов стенок под действием70 литических трансгликозилаз замедленна, что также наблюдается в эксперименте с Rpf,
представленном на рис.17.
Кроме этого, в пользу того, что Rpf является литическои трансгликозилазой, указывает наличие в его последовательности так называемого ES-фрагмента, который высоко консервативен и у белков семейства Rpf других микроорганизмов, и который характерен для активного центра литических трансгликозилаз из семейства 1 (рис. 20). Также для некоторых литических трансгликозилаз характерно наличие LysM домена, присутствующего и в структуре Rpf,
| Литичсская трансгл и ко з ил аза | Поел едоватсл ьность |
| LysG | —ES---------------------------------- GLMQV |
| Sit | RQES-F-P-A-S----------------- GAGLMQ |
| MltC | -TES-FNPYA-S---------------- A-GLMQV |
| EmtA | ATESGGNPNAVS------------ GLMQ |
| MltD | I VESA F- P - A- S------------ A-GLWQ |
| YfhD | YQESHWN P-A-S PTGVRGLMM |
 |
| 4SMDI 96 AYSV 50 GPSP 29G IKS 35 ATDO |
| Rv2450c RvlBMc RvTO Rv0667c |
| M.LUTE.RPF 47 ATVD |
Рис.20.
Последовательности некоторых литических "трансгликозилаз и белков семейства Rpf с указанием гомологичного ES-фрагмепта (Blackburn & Clarke, 2000). Следует отметить, что у литическои трапегликозилазы EmtA после ES-фрагмента идет последовательность GGN, то же самое имеет место у всех гомологов Rpf из М. tuberculosis.Действительно, препараты Rpf, в которых были проведены замены в ES-фрагменте и по двум цистеинам (Cys 50 на Lys, Cys ПО наТЬг), практически не оказывают литического действия на препараты клеток (Рис. 17).
71 В целом, можно предположить, что Rpf является белком, моделирующим состояние
клеточной стенки М, luteus, и ответственным за характер межклеточных взаимодействий в
культуре. Представляется, что его роль может быть двоякой: с одной стороны он может химически
модифицировать (вероятно» по механизму присущему литическим трансгликозилазам) какие-то
участки клеточной стенки, а с другой стороны играть структурную роль при формировании
межклеточных контактов, как это показано для других литических трапегликознлаз.
Образование клеточных агрегатов часто является ответом бактериальной популяции на
неблагоприятные условия окружающей среды. И если предположить, что Rpf участвует в
регуляции этого процесса, его роль представляется важной в переживании бактериями
неблагоприятных условий и последующем переходе к активному росту при наступлении условий,
способствующих дальнейшему росту микроорганизмов.
По данным С, Фостера среди белков, участвующих в модификации клеточной стенки
спорулирующих бактерий, обязательной для прорастания, спор, присутствуют и литическис
трансгликозилазы, очевидное сходство с которыми наблюдается у Rpf. Возможно, именно эта
ферментативная активность обуславливает способность Rpf реактивировать покоящиеся клетки
микрококка, например, способствуя локальному увеличению проницаемости клеточной стенки и
приводящему к активации клеточного метаболизма и деления, С другой стороны роль Rpf в
оживлении покоящихся клеток может состоять в диспергировании агрегатов клеток при их
реактивации.
72
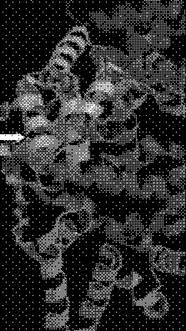
21 Трепгашя слрунтура укжшн лшоцимшодооный фолд Явжщнжтояабный фолд Rpf приведен для сравнения
73 3.4 Иммунологические свойства белков семейства Rpf
Наряду с изучением биохимических особенностей белка Rpf M. luteus, представляется важным выяснить, какими иммунологическими свойствами обладают белки семейства Rpf из
Micobacterium tuberculosis, каковы параметры иммунного ответа на них, и может ли иммунный ответ против этих белков быть протективным в отношении туберкулеза. Для решения данных задач были использованы методики и модели инфекции, разработанные нашими коллегами из Лаборатории иммунологии ЦНИИ туберкулеза РАМН, Представленные ниже эксперименты проводились совместно.
Поскольку белок Rpf играет роль фактора роста у MAuleus , можно предположить, что и микобактерии используют белки этого семейства для стимуляции деления. В пользу этого предположения свидетельствуют данные о том, что антитела к Rpf (реагирующие также и с Rpf-подобными белками М. tuberculosis) ингибируют рост микобактерии in vitro (Mukamolova et al, 2002}
Исходя из вышеприведенных фактов мы предположили, что предварительная иммунизация мышей рекомбинантными белками семейства Rpf могла бы индуцировать иммунный ответ, способный нейтрализовать биологическую активность соответствующих белков М. tuberculosis при заражении иммунизированных животных микобактериями и оказывать протективный эффект при экспериментальной туберкулезной инфекции,
Задачей данного раздела работы явилось изучение некоторых параметров иммунного ответа на Rpf-подобные белки микобактерии. В частности, была исследована продукция специфических антител и - поскольку основную роль в протективном иммунном ответе на М. tuberculosis играют Т-лимфоциты - пролиферативный ответ клеток лимфоузлов на Rpf-белки в тесте in vitro. Кроме того, были проведены исследования возможного протективного эффекта белков семейства Rpf в модели экспериментального туберкулеза на линейных мышах»
 В сыворотках с помощью иммуеофсрменшош теста исследовали уровень специфических пел класса BgG к белкам семейства Rpf (Рис 22).
В сыворотках с помощью иммуеофсрменшош теста исследовали уровень специфических пел класса BgG к белкам семейства Rpf (Рис 22).
22* Количество антител класса IgG к гомодошчньш Ярйштагетам в сынарт кях иммунных и интактаых мышеи {оптическая плотность, разведение сывороток
Как видно да результатов эксперимента, все белки семейства Rpf, за исключением Rpf t3, обладают высокой иммуио пзшюаъю и стимулируют образование антител в высоких
Вероятно, эта
сшшвд -
75
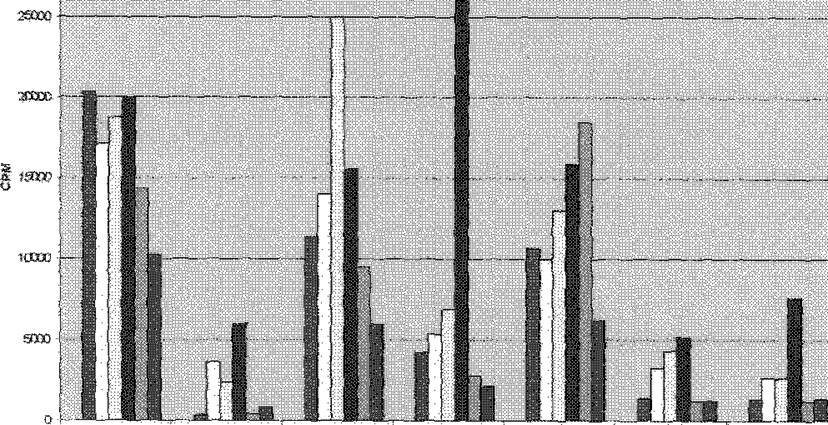
Уровень пршвдферааии оценмваяи да включению радкоакгавяой метки ь ДНК исследуемых клеток. Полученные результаты представлены на рис. 23
i£ t$ t4 is Rpf soplo emir
Pet, 25. Оценка проллф^ратиззного ответа клеток лвдфоузлов иммунных VE *шта#тдах мышей на тдалошчлыФ Rpf-айтяшш по жлютению ' Н-тишщиш.
Высокая шшуногеншеть белков Rpf ш vitro указывала на возможность протс&ттвдго эффекта йадилунм-зацйи щжушв туберкулеза в модели m гдю Д?я этот мышей линии 86 трижды иътувтир&в&дн 10 мкг соответствующего банка в штттм адъюв&ате ФроЙеда с интервалами в 2 недели, а затем ик заряжали легальной дозой M,№bem* fasti H37Rv,
Tlpii оценке ир^тгеггйвпой акхишосш белкоа с^мзйствд Kpf 8ш& исследована вмеев&емость M.htberctdosis из органов $аряжешня мышей. Цолуаднные результаты иредеяэдлшы в табл.3.
76
| Иммунизация | КОЕ на легкое |
| Контроль | 2.4х109±1.7х109 |
| Т2 | 5.2х108±1.4х108 |
| тз | 2.9х109+1.7х109 |
| Т4 | 2.8х108±1.4х108 |
| Т5 | 2.2х108±1.1х108 |
| Rpf | 2.0х 108± 1.1 хЮ8 |
Таблица 3. Высеваемость Mycohacterium tuberculosis из органов (количество КОЕ на орган) у мышей В6 через 3 недели после заражения.
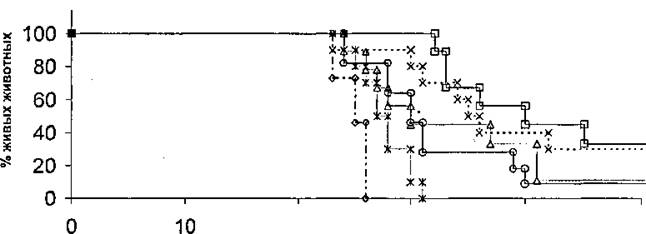
Как видно из приведенных результатов, иммунизация белками семейства Rpf снижала высеваемость микобактерий из легких не менее чем - на порядок по сравнению с неиммунизированным контролем. Следовательно, такая иммуниизация до некоторой степени защищает мышей от туберкулезной инфекции. Кроме того, протективный эффект иммунизации оценивался по продолжительности жизни иммунизированных животных относительно контроля (Рис.24),
| 120 i |
 77
77
| _,о-- | -control |
| -12 | |
| ___ _А_ | t3 |
| --•К- | -t4 |
| —ж- | -t5 |
| -П___ | -Rpf |
20 ' 30
Дни после заражения Рис.24. Динамика гибели вакцинированных мышей после внутривенного заражения
Mycobacterium tuberculosis.
Результаты опытов позволяют сделать вывод о том, что продолжительность жизни иммунизированных животных достоверно превышает таковую в контроле» В случае Rpf 12-14 у 40% иммунизированных мышей время жизни после заражения увеличивалось более чем в два раза. Таким образом, * полученные результаты свидетельствуют о том, что белки семейства Rpf обладают иммуногснными свойствами как in vitro, так и in vivor что позволяет рассматривать эти белки в качестве перспективных кандидатов на включение в состав субъединичных вакцин против туберкулеза,
В ходе исследования протективных свойств белков семейства Rpf была обнаружена одна чрезвычайно любопытная закономерность: протектнвные свойства белков оказались напрямую связанными со способом их хранения. На первом этапе провели два защитных эксперимента, причем в первом случае были получены результаты, свидетельствующие о значительном защитном эффекте белков семейства Rpf, а во втором результаты в опыте были на уровне контроля. При сравнении методик вакцинации в первом и во втором случаях выяснилось, что различие состояло лишь в том, что в случае протекции белки все время хранились при + 4 °С, а когда защитного эффекта не было, препараты белков хранились при температуре - 20 °С в глицерине. В качестве контрольного эксперимента был взят свежевыделенный препарат Rpf, который разделили на две части, хранившиеся, соответственно при + 4 °С и при - 20 °С, и иммунизировали ими две группы
78
мышей, а затем оценивали защитный эффект в обеих группах. D ходе этого исследования наши
предположения полностью подтвердились: в той группе животных, которая была иммунизирована белком, хранившимся при + 4 С, наблюдался существенный защитный эффект, в то же время, иммунизация животных белками, хранившимися при - 20 °С не приводила к защите.
Впоследствии для иммунизации использовались только белки, хранившиеся при + 4С, что неизменно приводило к протекции,
Возможно, подобное различие в иммунологических свойствах препаратов белка хранившихся в разных условиях может быть связано со склонностью белков семейства Rpf к агрегации. Как было показано выше Rpf чрезвычайно склонен к образованию агрегатов, особенно при длительном хранении (так при хранении в течении трех месяцев стерильного препарата белка при + 4 °С порядка 25 % белка переставали проходить через бактериальный фильтр). Некоторые из агрегатов очень велики и имеют молекулярную массу порядка, миллиона дальтон, а возможно и больше. В тоже время хранение препаратов при - 20 °С в 50 % глицерине препятствует образованию самых крупных агрегатов (не опубликованные наблюдения). В литературе, посвященной иммунному ответу на Л/ tuberculosis, встречаются данные о том, что белки, которые при иммунизации находятся в сильно агрегированном состоянии, в большей степени вызывают иммунный ответ клеточного типа, а именно этот тип ответа в основном защищает от туберкулеза (Sleytr et aL, 1997), Видимо, данный феномен можно объяснить именно различной степенью агрегации в препаратах белков, хранившихся в разных условиях. Не исключено, что такие свойства препаратов Rpf могут быть использованы в дальнейшем для создания протективных антитуберкулезных вакцин,
Заключение
В ходе дайной работы был изучен ряд свойств, присущих Rpf и его гомологам из М tuberculosis. До начала работы над этой диссертацией о белках семейства не было известно практически ничего, за исключением того факта» что Rpf M. luteus оказывает стимулирующее рост действие на покоящиеся клетки этого микроорганизма. Именно в силу этого были проведены
79
исследования в разных направлениях, целью которых являлось получение представлений о
механизме воздействия Rpf на клетки микрококка.
На первом этапе работы была разработана методика, позволяющая получать значительные количества рекомбинаптных белков семейства Rpf высокой чистоты. Эти белки использовались в дальнейших исследованиях, в частности, для иммунизации животных в защитных экспериментах на модели мышиного туберкулеза,
Также, при помощи нескольких методов, была изучена динамика накопления Rpf в культуре М, luteus при выращивании на различных средах, В целом, в ходе этого этапа работы удалось установить, что накопление Rpf в среде происходит на протяжении всего активного роста культуры, особенно на самых ранних его стадиях, однако в стационарной фазе синтез Rpi полностью прекращается и белок исчезает из среды. Эти данные позволила предположить, что активность изучаемого белка каким-то образом связана с делением клеток и ростом культуры- В то же время, наличие в структуре Rpf lys-M домена указывало на внеклеточный (поверхностный) характер его действия, В совокупности эти особенности организации Rpf привели нас к предположению об участии Rpf в процессах э связанных с метаболизмом клеточной стенки. Действительно, компьютерный анализ последовательности консервативного домена Rpf выявил наличие у него так называемого лизоцимо-подобного фолда, характерного для активного центра лизоцимов и литических трансгликозилаз - ферментов участвующих в гидролизе клеточной стенки бактерий. Обнаруженное разрушение крупных клеточных агрегатов М, luteus in vitro под действием Rpf могло также свидетельствовать в ползу гидролитической активности Rpf, В прямых экспериментах по воздействию Rpf на препарат лиофилизированных клеток М. luteus было получено еще одно доказательство литической активности. Наконец, данные сайт-направленного мутагенеза также подтверждают вывод о ферментативных свойствах изучаемого белка, В тоже время, поскольку замена глутамина- 13, в ES участке Rpf- известном фрагменте являющимся высоко консервативным для ряда трансгликозилаз из других объектов приводит к потере
80
литического действия, можно предположить, что белки семества Rpf относятся именно к этому
типу ферментов, осуществляющих процессинг клеточной стенки бактерий. { У
Изучение иммунологических свойств белков семейства Rpf в моделях туберкулеза на мышах выявило, что эти белки являются индукторами как гуморального так и клеточного ответов, а иммунизация ими животных приводит к протективному эффекту на модели экспериментального туберкулеза. Это позволяет рассматривать белки семейства Rpf в качестве перспективных кандидатов на включение в состав субъединичных вакцин против туберкулеза.
81
Еще по теме 3.3 Действие Rpf на клеточную стенку Л/, tuteus:
- 3- Ферменты клеточной стенки бактерий, участвующие в реактивации покоящихся форм
- Клеточно-молекулярные механизмы, лежащие в основе быстрого нейропротектирующего действия ИЛ-10 на мозг
- 3-2. Изучение синтеза Rpf и его гомологов М. tuberculosis в растущей культуре
- 4. Стенка
- 9.2 Метастазирование в стенку трахеи
- Место сосудистой стенки в понимании природа ГБ.
- Разрыв свободной стенки желудочка
- Утолщение стенки двенадцатиперстной кишки
- Утолщение стенки двенадцатиперстной кишки
- Оценка опухолевой инвазии в пределах кишечной стенки
- Предупреждающие действия — действия, предпринятые для устранения причины потенциального несоответствия.
- Казарьян, Константин Александрович. Биохимические и иммунологические свойства Белков семейства Rpf — факторов роста [Электронный ресурс] Micrococcus Iuteus и Mycobacterium tuberculosis : Дис. ... канд. Биол. наук : 03.00.04 .-М.: РГБ, 2003, 2003