Родина Picornaviridae
До складу родини Picornaviridae входять 12 родів.
1. Рід Aphtovirus - вірус ящуру (штами А, ASIA 1, С, О, SAT1-3).
2. Рід Avihepatovirus - вірус гепатиту А качок першого, другого, третього типів.
3. Рід Cardiovirus - віруси Тейлора, енцефаломіокардиту мишей (два серотипи).
4. Рід Enterovirus включає 10 видів: ентеровіруси ВРХ (ВРХ 1, 2); ентеровіруси людини групи А (віруси Коксакі А2-А8, А10, А12, А14, А16, ентеровіруси людини 71, 76, 89-92, 114, віруси мавп 19, 43, 46, ентеровірус бабуїнів А13); ентеровіруси людини групи В: віруси Коксакі В1-В6, А9, ЕСНО (від англ. enteric cytopathic human orphans) 1-7, 9, 11-21, 24-27, 29-33, ентеровіруси людини 69, 73-75, 77-88, 93, 97, 98, 100, 101, 106, 107, 110, мавп'ячий агент 5; ентеровіруси людини групи С (поліовіруси 1-3, віруси Коксакі А1, А11, А13, А17, А19-А22, А24, ентеровіруси людини 95, 96, 99, 102, 104, 105, 109, 113, 116); ентеровіруси людини групи D: 68, 70, 94, 111; риновіруси людини групи А: А1, 2, 7, 8-13, 15, 16, 18-25, 28-34, 36, 38-41, 43-47, 49-51, 53-68, 71, 73-78, 80-82, 85, 88-90, 94-96, 98, 100-103; риновіруси людини групи В: В3-6, 14, 17, 26, 27, 35, 37, 42, 48, 52, 69, 70, 72, 79, 83, 84, 86, 91-93, 97, 99; риновіруси людини групи С: С1-49; ентеровіруси свиней групи В: В9, В10; ентеро- віруси мавп групи А: А1.
5. Рід Erbovirus - вірус інфекційного риніту коней (два серотипи).
6. Рід Hepatovirus - вірус гепатиту А людини (раніше ентеровірус людини типу 72), вірус гепатиту А мавп.
7. Рід Kobuvirus - вірус Аїчі (Aichi virus) - викликає гастроентерити в людей і кобувірус корів.
8. Рід Paraechovirus - людські параеховіруси (чотири серотипи).
9. Рід Sapelovirus - ентеровірус свиней А (Porcine sapelovirus), 8У2-подібні віруси (Simian sapelovirus) і пікорнаві- рус качок TW90A (Avian sapelovirus).
10. Рід Senecavirus - вірус долини Сенека (свиней).
11. Рід Teschovirus - вірус хвороби Тешена.
12. Рід Tremovirus - вірус енцефаломієліту птахів.
Віріони ікосаедричні, без оболонки (рис. 1.17). Діаметр часток 22-30 нм. Серцевина складається з одноланцюгової РНК і малого білка 3BVpg, що ковалентно приєднаний до її 5'-кінця. При лужному рН іноді формуються видовжені рибонуклеопро- теїнові структури. Капсид складається з 60 білкових субодиниць (попередник - продукт гена Р1), а кожна з них - з чотирьох білків: трьох із масою від 32 до 41 кДа (VP1-VP3) та одного з масою 5,5-13,5 кДа (VP4).
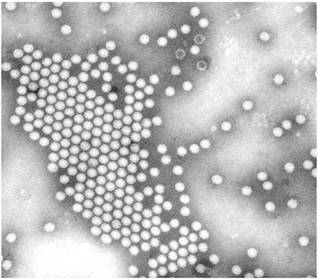
Рис. 1.17. Електронномікроскопічне зображення часток вірусу поліомієліту [50]
Віріон містить одну молекулу лінійної олРНК позитивної полярності. Розмір 7-8,5 тис. о. На З'-кінці розміщений полі(А), його довжина залежить від роду. Малий білок VPg (М = 24 кДа) ковалентно приєднаний до 5'-кінця. Некодуюча ділянка 5'-кінця має визначену вторинну структуру, що є необхідною для синтезу ланцюгів РНК негативної полярності, поліаденілування, інфекційності; бере участь у приєднанні VpgUUU. Окрім того, на 5'-кінці є //?/А-структура (Internal Ribosome Entry Site). За некодуючою послідовністю на 5'-кінці розташована відкрита рамка зчитування білка-попередника (М = 240-250 кДа) структурних білків (Р1) і переважно неструктурні білки (Р2, РЗ), за якими розташовані коротка некодуюча послідовність і полі(А). У деяких вірусів послідовностям структурних білків передує лідируюча послідовність (L). Ідентичність послідовностей між вірусами різних родів становить 40 %.
Білок-попередник нарізається на функціональні білки протеазами. До неструктурних білків, що мають протеолітичну активність, відносять Lpro, 2Apro, 3Cpro; інші неструктурні білки містять полімеразу (3Dpol), АТФазу (2C) і білок з невідомою функцією (2А, 2В). Лідируючий білок афтовірусів має протеолітичну активність (Lpro), хоча в карідовірусів такої активності немає.
Проміжні форми при протеолітичному нарізанні можуть також виявляти певні функції (наприклад, протеолітична активність асоціюється з проміжним продуктом 3CD поліовірусів).Капсиди сформовані з 60 ідентичних субодиниць (протоме- рів), кожна з яких складається з чотирьох поверхневих білків - продуктів гена P1 (1A, 1B, 1C, 1D) у поліовірусів VP4, VP2, VP3, VP1 та однієї копії білка, зв'язаного з геномом 3bVpg. Крім того, попередник білків VP4 та VP2 (VP0) також може бути в структурі капсиду.
Для віріонів не характерна наявність ліпідів. Проте в деяких штамів поліовірусів у структурі віріонів може бути до 60 сфингозиноподібних молекул. Поліпептид 1А (VP4), що є у внутрішній поверхні капсиду, має молекулу міристичної кислоти, яка ковалентно приєднана до амінокінця гліцину.
Вірусні білки не глікозильовані.
Нативні віріони мають подібні типоспецифічні антигени (N чи D), які після легкого нагрівання перетворюються на групоспецифічні антигени (Н чи С). Серотипи вірусів класифікуються на основі реакції перехресної нейтралізації інфекційності, зв'язування комплементу, специфічного імуноферментного аналізу та реакції імунодифузії.
Маса віріонів 8-9 х 106 кДа. Густина в градієнті CsCl 1,33-1,45 г/см3. Sw20=140-165 S. Деякі частки нестабільні при рНщо викликає рак шкіри Меркеля, а в 2010 р. - поліомавірус, що призводить до тиходисплазії (Trichodysplasia spinulosa virus, TSV) - рідкісного захворювання шкіри, що виникає під час імуносупресивної терапії в пацієнтів із трансплантаціями.
Еще по теме Родина Picornaviridae:
- Семейство Пикорнавирусов (Family Picornaviridae)
- Родина Orthomyxoviridae
- Родина Coronaviridae
- Родина Papillomaviridae
- Родина Iridoviridae
- Родина Asfaviridae
- Родина Bunyaviridae
- Родина Togaviridae
- Родина Reoviridae
- Родина Parvoviridae
- Родина Caliciviridae
- Родина Arenaviridae
- Родина Flaviviridae
- Родина Herpesviridae
- Родина Hepadnaviridae
- Родина Rhabdoviridae