Роль тау-белка в развитии БА
Нейрофибриллярные клубки - это нитевидные включения, которые накапливаются в значительном числе нейронов мозга пациентов с БА. Данные патологические включения также встречаются и при других нейродегенеративных расстройствах, включая фронтотемпоральную деменцию с паркинсонизмом, сцепленную с 17-й хромосомой (Frontotemporal dementia with parkinsonism-17, FTDP-17), болезнь Пика, прогрессивный супрануклеарный паралич (Progressive Supranuclear Palsy, PSP) и кортикобазальную дегенерацию (Corticobasal degeneration, CBD).
Основным компонентом нейрофибриллярных клубков является белок тау, ассоциированный с микротрубочками (Goedert et al., 1988; Grundke-Iqbal et al., 1986a). В нормальном состоянии тау представляет собой растворимый белок, который способствует сборке и стабилизации микротрубочек. При гиперфосфорилировании тау-белка он переходит в патологическое состояние с последующим формированием нитевидных структур (рис. 2) (Goedert et al., 1992; Grundke-Iqbal et al., 1986b).Было показано, что гиперфосфорилированный тау проявляет сниженное сродство к микротрубочкам (Biernat et al., 1993; Bramblett et al., 1993). Белок тау кодируется в геноме мыши и человека единственным геном (MAPT), расположенным на 17-й хромосоме, а в результате альтернативного сплайсинга в мозге взрослого человека образуются шесть основных изоформ белка (Goedert et al., 1989). Ген MAPT содержит 15 экзонов; 2-й, 3-й и 10-й могут подвергаться альтернативному сплай-
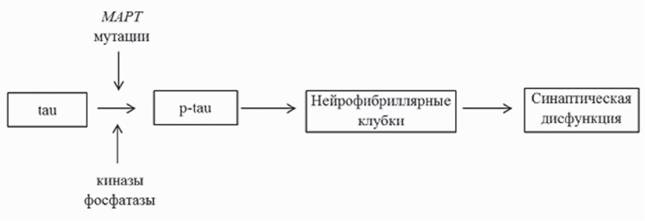
Рис. 2. Роль тау-белка в патогенезе БА в рамках тау-гипотезы
сингу. Четыре несовершенных тандемных повтора кодируются экзонами 9-12, поэтому альтернативный сплайсинг 10-го экзона дает изоформы, содержащие по три или по четыре домена с повторами, обозначаемые как 3R- и 4R-формы тау, в зависимости от того, отсутствует или присутствует 10-й экзон, соответственно.
Альтернативный сплайсинг экзонов 2 и 3 дает варианты, содержащие ноль (0N), одну (1N) или две (2N) вставки на N-конце, так что формируется шесть изоформ тау: 3R0N, 3R1N, 3R2N, 4R0N, 4R1N и 4R2N. В мозге взрослого человека соотношение 3R к 4R тау составляет 1:1, в то время как в мозге взрослых мышей тау представлен единственной изоформой 4R (Gotz et al., 1995; Janke et al., 1999). Таупатии могут быть дополнительно классифицированы в зависимости от того, состоят ли клубки из 3R- или 4R-изоформы. Например, при БА оба варианта (3R и 4R) тау накапливаются в нейрофибриллярных клубках, при других заболеваниях отмечаются только 3R- (например болезнь Пика) или 4R-формы тау (например при прогрессирующем супрануклеарном параличе и кортикобазальной дегенерации) (Delacourte et al., 1998; Sergeant et al., 1999). При БА тау-патология наблюдается только в нейронах, однако при других тау-патиях, таких как 4R тау-патии, характерные для прогрессирующего супрануклеарного паралича и кортикобазальной дегенерации, тау-включения наблюдаются также и в глии (Lee et al., 2001). Ген, кодирующий тау, генетически не ассоциирован с БА, а выявленные в нем мутации ассоциированы с некоторыми формами фронтотемпоральной дегенерации (ФТД) (Spillantini et al., 1998). В результате идентификации патогенных мутаций в гене, кодирующем тау (MAPT), было показано, что дисфункции тау самой по себе достаточно для того, чтобы вызвать нейродегенерацию (Hutton et al., 1998). Однако отсутствие генетической связи с БА указывает на то, что тау-патология в каскаде процессов нейродегенерации находится после развития амилоидоза мозга. Это не означает, что тау-патология не имеет значения в патогенезе БА, более того, нейродегенерация, вызванная дисфункцией тау, может играть центральную роль при БА. Возможно, что тау-патология может быть вызвана различными механизмами - как зависящими от Ap, так и не зависящими от нарушения процессинга АРР.1.
Еще по теме Роль тау-белка в развитии БА:
- Агрегация РНК-связывающего белка FUS как патогенетический фактор развития нейродегенеративного процесса
- Взаимосвязь между Ap и тау
- 2. Понятие общения и его роль в развитии ребенка. Смена форм общения ребенка со взрослым от рождения до 7 лет в Норме и при отклонениях в развитии
- Тау-белки нейронов
- Роль инфекции в развитии СРК
- 1.8 Роль хламидийной инфекции в развитии сочетанной патологии
- Роль цитокинов и других гуморальных факторов в развитии кахексии.
- 58.Роль раннего детства в формировании и развитии одарённости.
- Роль тестостерона и дигидротестостерона в развитии ДГПЖ
- 27. Роль наследственности и среды в формировании и развитии психофизиологических функций.
- 9. Социальные причины дефектов и их роль в развитии аномальных детей.
- Роль лимфатической системы в развитии отека
- 8. Биологические причины дефектов и их роль в развитии аномальных детей.
- 42. Общение, игра, обучение и труд и их роль в развитии человека.
- Роль аутоаллергических (аутоиммунных) механизмов в развитии эндокринных нарушений
- Роль наследственных факторов в возникновении и развитии туберкулеза
- 1.5 Роль воспаления в развитии ишемической болезни сердца
- Образование нейрофибриллярных сплетений из гиперфосфорилированных тау-белков
- Структура индивидных свойств человека и их роль в развитии личности.
- 1.3 Роль микробного фактора в развитии хронического генерализованного пародонтита