нАПЮГНБЮМХЕ МЕИПНТХАПХККЪПМШУ ЯОКЕРЕМХИ ХГ ЦХОЕПТНЯТНПХКХПНБЮММШУ РЮС-АЕКЙНБ
оПХ ХЯЯКЕДНБЮМХХ ЛНГЦЮ ОЮЖХЕМРНБ, СЛЕПЬХУ НР аю, ЙПНЛЕ ЯЕМХКЭМШУ АКЪЬЕЙ НАШВМН НАМЮПСФХБЮЧР ХМРПЮМЕИПНМЮКЭМШЕ МЕИПНТХАПХККЪПМШЕ ЯОКЕРЕМХЪ (хмтя) (Avila et al., 2004; Johnson, Bailey, 2002).
оПХ ЩКЕЙРПНММНИ ЛХЙПНЯЙНОХХ НМХ ХЛЕЧР БХД ОЮПМШУ ЯОХПЮКЭМН ЯЙПСВЕММШУ МЕИПНТХКЮЛЕМРНБ Б ЖХРНОКЮГЛЕ МЕИПНМНБ, ЙНРНПШЕ ЯНЯРНЪР ХГ ЦХОЕПТНЯТНПХККХПНБЮММШУ ра. оПХ ЩРНЛ НДХМ ТНЯТЮР ОПХЯНЕДХМЪЕРЯЪ Й НДМНИ ЛНКЕЙСКЕ ра, Ю БРНПНИ ТНЯТЮР ОНРЕМЖХЮКЭМН ЦНРНБ БЯРСОХРЭ Б ПЕЮЙЖХЧ Я ДПСЦНИ ЛНКЕЙСКНИ ра. оПХ НОПЕДЕКЕММНИ ЯХРСЮЖХХ ДПСЦЮЪ ЛНКЕЙСКЮ ра ОПХЯНЕДХМЪЕР ЩРНР ТНЯТЮР Х НАПЮГСЕРЯЪ ДХЛЕП, ЯНЕДХМЕММШИ ТНЯТЮРМШЛ ЛНЯРХЙНЛ (Rao et al., 2009).кНЦХВМЮЪ ЦХОНРЕГЮ Н ПНКХ ТНЯТНПХКХПНБЮМХЪ Б ТНПЛХПНБЮМХХ хмтя ХГ ра АШКЮ ЯТНПЛСКХПНБЮМЮ Б 2005 Ц. (Iqbal, Grundke-Iqbal, 2005). б ЯННРБЕРЯРБХХ Я ОПЕДЯРЮБКЕМХЪЛХ ЩРХУ ЮБРНПНБ, ЛНКЕЙСКШ ра НАПЮГСЧР ЦНЛНЮЦПЕЦЮРШ, Б НЯМНБМНЛ ОНЯПЕДЯРБНЛ ДНЛЕМНБ, ЙНРНПШЕ БГЮХЛНДЕИЯРБСЧР Я РСАСКХМНЛ ЛХЙПНРПСАНВЕЙ (ПХЯ. 4).
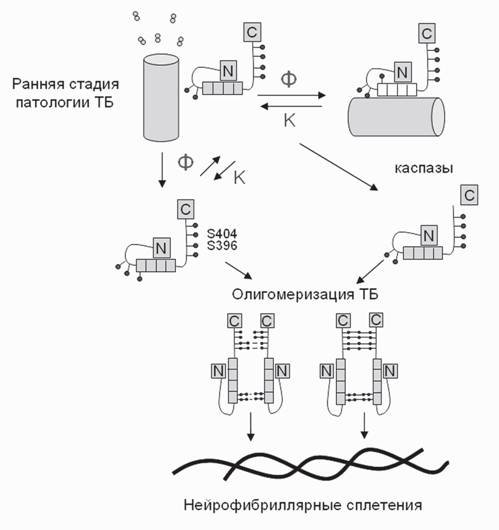
пХЯ. 4. яУЕЛЮ СВЮЯРХЪ ЯЮИР-ЯОЕЖХТХВЕЯЙНЦНЦХОЕПТНЯТНПХКХПНБЮМХЪ (Ser404 Х Ser396) Х ЙЮЯОЮГ- МНЦН ПЮЯЫЕОКЕМХЪ ра Б ОПНЖЕЯЯЕ ЮЦПЕЦЮЖХХ Я НАПЮГНБЮМХЕЛ МЕИПНТХАПХККЪПМШУ ЯОКЕРЕМХИ й - ЙХМЮГШ, т - ТНЯТaРЮГШ, N- Х C- - ЙНМЖШ ЛНКЕЙСКШ ра. жХКХМДП - ЛХЙПНРПСАНВЙЮ
цХОЕПТНЯТНПХКХПНБЮМХЕ ра МЕИРПЮКХГСЕР ДЮММШЕ ДНЛЕМШ, НАКЕЦВЮЪ ЮЦПЕЦЮЖХЧ Х ТНПЛХПНБЮМХЕ хмтя. щЙЯОЕПХЛЕМРЮКЭМН АШКН ДНЙЮГЮМН, ВРН ТНЯТНПХКХПНБЮМХЕ ра ОН ЯЮИРЮЛ рзЦ231 Х Ser262 ЯСЫЕЯРБЕММН ЯМХФЮЕР ЕЦН БГЮХЛНДЕИЯРБХЕ Я РСАСКХМНЛ ЛХЙПНРПСАНВЕЙ (Biernat et al., 1993; Cripps et al., 2006). мЕЙНРНПШЕ ЮБРНПШ ЯВХРЮЧР ТНЯТНПХКХПНБЮМХЕ МЕНАУНДХЛШЛ, МН МЕДНЯРЮРНВМШЛ СЯКНБХЕЛ ДКЪ ОНКХЛЕПХГЮЖХХ ра (Chun, Johnson, 2007a,b). яЮИР-ЯОЕЖХТХВЕЯЙНЕ ТНЯТНПХКХПНБЮМХЕ ра ЯНОПНБНФДЮЕРЯЪ ЕЦН ПЮЯЫЕОКЕМХЕЛ ЙЮЯОЮГНИ-3 ОН ЯЮИРЮЛ Ser422 Х Ser396/404 (Ferrari et al., 2003; Haase et al., 2004). сЙНПНВЕММШЕ Х ЦХОЕПТНЯТНПХКХПНБЮММШЕ ЛНКЕЙСКШ ЩРНЦН АЕКЙЮ ЯСЫЕЯРБЕММН ЮЙРХБМЕЕ ТНПЛХПСЧР МЕИПНТХАПХККЪПМШЕ ЯОКЕРЕМХЪ.
нОХЯЮММНЕ ПЮЯЫЕОКЕМХЕ АЕКЙЮ ОН Glu391 Х ДЕЮЛХДХПНБЮМХЕ ОН юй- НЯРЮРЙЮЛ ЮЯОЮПЮЦХМЮ (ХКХ ЦКСРЮЛХМЮ) РЮЙФЕ ЯОНЯНАЯРБСЧР ЮЦПЕЦЮЖХХ (Ding et al., 2006). рНЙЯХВМНЯРЭ ЦХОЕПТНЯТНПХКХПНБЮММНЦН ра, ОН ЮМЮКНЦХХ Я юП, ЮЯЯНЖХХПСЕР Я ЙНПНРЙХЛХ НКХЦНЛЕПЮЛХ Х МЕАНКЭЬХЛХ ЮЦПЕЦЮРЮЛХ, Ю МЕ Я ЙПСОМШЛХ МЕИПНТХКЮ- ЛЕМРЮЛХ (Spires-Jones et al., 2008). мЮ МЕИПНМЮУ ЛШЬЕИ АШКН ОПНДЕЛНМЯРПХПНБЮМН НРЯСРЯРБХЕ РНЙЯХВЕЯЙНЦН ЩТТЕЙРЮ ТюП Б СЯКНБХЪУ МНЙЮСРЮ Р-ЦЕМЮ Х МЕИПНРНЙЯХВМНЯРЭ ОЯЕБДНТНЯТНПХКХПНБЮММНЦН ЩЙГНЦЕММНЦН ра (Fath et al., 2002; Rapoport et al., 2002). оН МЮЬЕЛС ЛМЕМХЧ, ДНАЮБКЕМХЕ ТюП Б СЯКНБХЪУ МНЙЮСРЮ Р-ЦЕМЮ МЕ БШГШБЮЕР ЙНККЮОЯЮ, РЮЙ ЙЮЙ ТюП ЯРХЛСКХПСЕР ЯХМРЕГ АЕКЙЮ Б МЕИПНМЮУ, С ЙНРНПШУ МЕР ра, ОНЩРНЛС МЕ ОПНХЯУНДХР ЦХОЕПТНЯТНПХКХПНБЮМХЪ ра Х ОНЯКЕДСЧЫЕЦН ЙНККЮОЯЮ. сАЕДХРЕКЭМН ОНЙЮГЮМН, ВРН ОПХ ХГАШРНВМНИ ЩЙЯОПЕЯЯХХ ра МЮПСЬЮЕРЯЪ ЮЙЯНМЮКЭМШИ РПЮМЯОНПР Б МЕИПНМЮУ. сЦМЕРЮЧРЯЪ АШЯРПШИ ЙХМЕГХМГЮБХЯХЛШИ ЮМРЕПНЦПЮДМШИ РПЮМЯОНПР Б ЯБЪГХ Я АКНЙНЛ ЛХЙПНРПСАНВЕЙ Х ЛЕДКЕММШИ ПЕРПНЦПЮДМШИ РПЮМЯОНПР, ОНЯЙНКЭЙС МЕИПНТХКЮЛЕМРШ РЪФЕКШУ ЖЕОЕИ Х ра ЙНМЙСПХПСЧР ГЮ БГЮХЛНДЕИЯРБХЕ Я я-ЙНМЖЕБШЛ ТПЮЦЛЕМРНЛ РСАСКХМЮ (Ebneth et al., 1998; Probst et al., 2000). ра РПЮМЯОНПРХПСЧРЯЪ Б НЯМНБМНЛ ЛЕДКЕММШЛ ЮЙЯНМЮКЭМШЛ ОСРЕЛ, ОПХ ЩРНЛ ХУ ХГАШРНЙ МЮПСЬЮЕР ЯБЪГЭ ЛХЙПНРПСАНВЕЙ Х МЕИПНТХКЮЛЕМРНБ, ОПХБНДЪ Й МЮЙНОКЕМХЧ ОНЯКЕДМХУ Б ЮЙЯНМЕ. нВЕМЭ ХМРЕПЕЯМШЕ ПЕГСКЭРЮРШ АШКХ ОНКСВЕМШ АНКЭЬНИ ЦПСООНИ ХЯЯКЕДНБЮРЕКЕИ МЮ ЙСКЭРСПЮУ МЕИПНМНБ, МНЙЮСРМШУ ОН ра (Tau-/-) Х МЕИПНМЮУ ДХЙНЦН РХОЮ (Tau+/+) (Vossel et al., 2010). юБРНПШ ХЯЯКЕДНБЮКХ РПЮМЯОНПР ЛХРНУНМДПХИ Х TrkA-ПЕЖЕОРНПНБ НР ЯХМЮОЯЮ Й РЕКС ЙКЕРЙХ (ПЕРПНЦПЮДМНЕ ДБХФЕМХЕ) Х НР РЕКЮ ЙКЕРЙХ Й ЯХМЮОЯС (ЮМРЕПНЦПЮДМНЕ ДБХФЕМХЕ). сЯРЮМНБКЕМН, ВРН ОПХАЮБКЕМХЕ Ap ╚БШГШБЮЕР РПЮМЯОНПРМШИ ЙНККЮОЯ╩ С Tau+/+ МЕИПНМНБ ДХЙНЦН РХОЮ (ЯНДЕПФЮЫХУ ра), МН МЕ С Tau-/- МЕИПНМНБ, МЕ ЯНДЕПФЮЫХУ ра. юБРНПШ МЕ НАЯСФДЮЧР, ЙЮЙХЛ НАПЮГНЛ НЯСЫЕЯРБКЪЕРЯЪ ЩРЮ ЯБЪГЭ, ЕЯКХ Ap МЕ СВЮЯРБСЕР Б ОПНЖЕЯЯЮУ ТНЯТНПХКХПНБЮМХЪ АЕКЙНБ. б ДЮММНИ ЯХРСЮЖХХ ДНАЮБКЕМХЕ Ap БШГШБЮЕР ХМРЕМЯХБМШИ ЯХМРЕГ АЕКЙЮ Б МЕИПНМЮУ. оНЩРНЛС Б МЕИПНМЮУ ДХЙНЦН РХОЮ ра МЮВХМЮЧР ЮЙРХБМН ТНЯТНПХКХПН- БЮРЭЯЪ Б ГНМЕ ЯХМРЕГЮ АЕКЙЮ Х ОПХ ДКХРЕКЭМНЛ БНГДЕИЯРБХХ Ap Б МЕИПНМЮУ МЮВХМЮЕР ПЮГБХБЮРЭЯЪ ОПНЖЕЯЯ ЦХОЕПТНЯТНПХКХПНБЮМХЪ, Ю ГЮРЕЛ РПЮМЯОНПРМШИ ЙНККЮОЯ.13.
еЫЕ ОН РЕЛЕ нАПЮГНБЮМХЕ МЕИПНТХАПХККЪПМШУ ЯОКЕРЕМХИ ХГ ЦХОЕПТНЯТНПХКХПНБЮММШУ РЮС-АЕКЙНБ:
- мЮПСЬЕМХЪ НАПЮГНБЮМХЪ Х БШБЕДЕМХЪ ЙНМЕВМШУ ОПНДСЙРНБ АЕКЙНБНЦН НАЛЕМЮ
- пНКЭ РЮС-АЕКЙЮ Б ПЮГБХРХХ аю
- бГЮХЛНЯБЪГЭ ЛЕФДС Ap Х РЮС
- рЮС-АЕКЙХ МЕИПНМНБ
- нОСУНКХ ЯНЯСДХЯРШУ ЯОКЕРЕМХИ
- йЮПЖХМНЛЮ ОЮСРХММНЦН ЯОКЕРЕМХЪ
- оНКМШИ ОЮПЮКХВ ОКЕВЕБНЦН ЯОКЕРЕМХЪ
- мЕБПХР ОКЕВЕБНЦН ЯОКЕРЕМХЪ.
- мХФМХИ ОЮПЮКХВ ОКЕВЕБНЦН ЯОКЕРЕМХЪ
- мЕБПЮКЦХЪ ОКЕВЕБНЦН ЯОКЕРЕМХЪ
- мЕБПЮКЦХЪ ЦКНРНВМНЦН ЯОКЕРЕМХЪ мЕБПЮКЦХЪ БЕПУМЕЦН ЦНПРЮММНЦН МЕПБЮ
- 1.4 яНБПЕЛЕММШЕ ЛЕРНДШ КЕВЕМХЪ ЙХЯРНГМШУ НАПЮГНБЮМХИ ЪХВМХЙНБ 1.4.1 уХПСПЦХВЕЯЙНЕ КЕВЕМХЕ НАПЮГНБЮМХИ ЪХВМХЙНБ
- мЮПСЬЕМХЕ НАЛЕМЮ АЕКЙНБ
- мЮПСЬЕМХЪ АЕКЙНБНЦН ЯНЯРЮБЮ ЙПНБХ