Тау-белки нейронов
Другим, наиболее активным и эффективным эвакуатором фосфатов и, возможно, пирофосфатов из зоны синтеза белка является ТБ. Белковая молекула ТБ является гидрофильным и высоко асимметричным белком.
N-концевой фрагмент ТБ взаимодействует с плазматической мембраной и цитоскелетными белками нейрона, принимает участие в сигнальной трансдукции, определяет расстояние между микротрубочками в аксоне и способен регулировать диаметр последнего (Brandt et al., 1995; Chen et al., 1992).Данный фрагмент содержит кислую и богатую пролином области в своем составе. Именно богатая пролином область ТБ интенсивно фосфорилирована в мозге пациентов с БА (Johnson, Jenkins, 1999). В составе N-концевого фрагмента ТБ есть небольшие участки связывания железа и гепарина. С-концевой фрагмент ТБ содержит 3 или 4 тубулин-связывающих домена длиной 31-32 остатка каждый (R) и один кислый регион (Alonso et al., 2006) (рис. 3). Установлено, что при активации гепарином GSK3P усиливается фосфорилирование ТБ по сайтам Ser199, Thr231, Ser235, Ser262, Ser396, Ser400, которые характерны для гиперфосфорилирован- ного ТБ при БА (Song, Yang, 1995). Ненасыщенные жирные кислоты, входящие в состав фосфолипидов, и ионы металлов, особенно алюминий, также стимулируют полимеризацию ТБ (Scott et al., 1993; Wilson, Binder, 1997). В настоящее время рассматривается гипотеза о роли так называемых «воспламеняющих» сайтов фосфорилирования ТБ (Johnson, Bailey, 2002). Речь идет о сайтах, фосфорилирование которых облегчает дальнейшее множественное фосфорилирование молекулы ТБ по аномальным сайтам с образованием гиперфосфорилированного белка. Как правило, «воспламеняющие» сайты в ТБ являются участками взаимодействия с другими внутринейрональными протеинами, и прежде всего тубулином, которому отводится ключевая роль в липид-белковых комплексах возбудимых мембран (Шноль, 1979). Таким образом, тубулин не только входит в состав трубочек, но является одним из компонентов в составе протоплазмы нейрона, определяющих свойства и характеристики фазового перехода возбудимых мембран.
К процессам, принимающим участие в посттрансляционной модификации ТБ, относятся фосфорилирование, убиквитирование, гликирование, гликозилирование, окисление, нитрование, агрегация и др. (Cripps et al., 2006; Kuhla et al., 2007). В норме ТБ, взаимодействуя с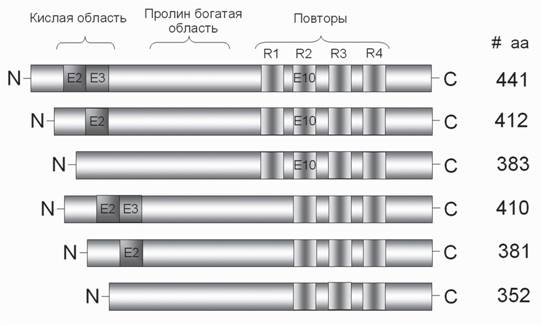
Рис. 3. Схема доменной структуры 6 изоформ тау-белков. Е2, Е3 и Е10 - экзоны N-концевого и С-концевого фрагментов белка
Повторы R1-R4 в молекулах ТБ, содержащие дополнительные сайты, способны связывать фосфаты и АМФ для транспорта из зоны синтеза белка к митохондриям. # аа - число аминокислотных остатков в изоформах ТБ
тубулином, скрепляет и стабилизирует микротрубочки (Biemat et al., 1993), обеспечивает перенос в цитоплазме нейронов клеточных органелл и мобильных биохимических молекул (Probst et al., 2000), регулирует рост аксонов и дендритов (Dawson et al., 2001). Способность ТБ скреплять микротрубочки отчасти объясняется наличием у него нескольких фосфатных групп (см. рис. 3). При интенсивном синтезе белка в нейроне в зоне синтеза белка образуется большое количество молекул пирофосфата и АМР. Пирофосфат может подвергаться гидролизу пирофосфатазой с образованием двух молекул фосфата, а выделяющаяся при этом энергия рассеивается в виде тепла. Молекулы ТБ при участии киназ присоединяют молекулы фосфатов в зоне синтеза белка и способствуют эвакуации этих фосфатов к митохондриям. Возможно, что аналогично осуществляется транспорт молекул АМР из зоны синтеза белка к митохондриям для восстановления в АТР. При продолжительном интенсивном синтезе белка в зоне синтеза температура может повышаться до определенного значения, при котором возникает вероятность присоединения нескольких молекул фосфатов к различным сайтам молекулы ТБ. Аксоногенез также регулируется уровнем фосфорилирования ТБ, при этом образуется градиент степени фосфорилирования молекул ТБ вдоль растущего аксона. Было убедительно показано, что в теле нейрона в основании аксона ТБ фосфорилирован по Ser199/202 и Thr205 на 80%, тогда как в конусе роста аксона только на 20% (Mandell, Banker, 1996).
Эти результаты четко свидетельствуют о том, что в центральных зонах нейрона молекулы ТБ поли- фосфорилируются, а затем, по мере перемещения ТБ от центра нейрона, фосфаты утилизируются. Процесс блокирования пирофосфатов имеет для нейрона важное значение, так как изымает из энергетического обращения энергоемкие молекулы и предотвращает повышение температуры в нейронах. Таким образом, формулу полифосфорилирования ТБ можно записать в виде:ТБ+nATP ~ ТБпР+nADP.
В ряде обзоров описаны варианты предотвращения гидролиза пирофосфатов пирофосфатазой в разных организмах (Rao et al., 2009; Da Silva et al., 1991; Terkeltaub, 2001). В нейронах, по нашему мнению, этот процесс протекает следующим образом:
nADP+nPPi ^ nATP+nPi.
Огромную роль в изучении патогенеза БА занимают исследования генетики ТБ. т-Ген располагается на 17 хромосоме (17q21 локус) и содержит по крайней мере 16 экзонов (Andreadis et al., 1992; Goedert et al., 1999). В настоящее время для т-гена выявлен полиморфный динуклеотидный повтор в 9 интроне и 8 одинарных нуклеотидных полиморфизмов (Baker et al., 1999; Rizzu et al., 1999). Первичный эффект интронных мутаций т-гена, находящихся рядом со сплайс-донорским участком, обусловлен изменением соотношения изоформ ТБ с различным количеством повторов, при этом имеет место преобладание изоформ с 4 повторами (Hong et al., 1998; Ryoo et al., 2007). Была выявлена роль еще одного фермента в гиперфосфорилировании ТБ. Речь идет о тирозин фосфорилирующей регуляторной киназе 1А с двойной специфичностью (DYRK1A). Ген DYRK1A локализован на той же 21 хромосоме, что и ген АРР. С этим ассоциирует наличие интранейрональных нейрофибриллярных сплетений (ИНФС) одновременно с внеклеточными сенильными бляшками агрегированного Ар при трисомии 21 хромосомы (у пациентов с синдромом Дауна), либо при увеличении копий отдельных генов данной хромосомы (болезнь Альцгеймера) (Grundke-Iqbal et al., 1986; Spires et al., 2006). В мозге человека идентифицировано 6 изоформ ТБ (45-65 кДа) длиной 352-441 а.к.
остатков, экспрессирующихся в результате альтернативного мРНК-сплайсинга 2-, 3- и 10-го экзонов (см. рис. 3). Изоформы ТБ отличаются наличием трех (R2, R3, R4) или четырех (R1, R2, R3, R4) повторяющихся участков в С-концевом фрагменте молекулы белка и наличием либо отсутствием одной из двух вставок (E2 и Е3) в N-концевом фрагменте. Для ТБ характерна биохимическая гетерогенность в различных субпопуляциях нейронов (Buee et al., 2000). Самая длинная изоформа ТБ (441 АК) содержит 80 остатков серина и 5 остатков тирозина, поэтому 20% молекулы потенциально может быть фосфорилировано (Goedert et al., 1989). Изоформы ТБ при разных заболеваниях регистрируются в разных сочетаниях. В частности, при БА идентифицирован триплет ТБ (изоформы тау 55, 64 и 69 кДа), являющийся основной составляющей ИФНС. Все они образуются в результате гиперфосфорилирования. В норме для различных изоформ ТБ выявлены специфические участки фосфорилирования. Однако при БА наблюдается неспецифическое гиперфосфорилирование целого ряда изоформ ТБ по аномальным сайтам (Kopke et al., 1993). Изоформы ТБ с экзоном 10 содержат 2 цистеиновых остатка, без этого экзона - только один. Это предполагает возможность формирования димеров белка за счет образования межмолекулярных дисульфидных связей. Возможно, что окисление сульфгидрильных групп в данном белке может вносить свой вклад в его агрегацию. В нейрофибриллярных сплетениях при БА обнаружены все 6 изоформ гиперфосфорилированного ТБ, которые при электрофоретическом разделении дают 3 основные полосы, соответствующие белкам с молекулярным весом 68, 64 и 60 кДа (Delacourte et al., 1999). Недавно обнаружена изоформа ТБ, представленная в ядре нейронов и других клеток. Она колокализуется с ядерным белком нуклеолином и АТ-богатым участком сателитной ДНК и представлена в конститутивном хроматине. Авторы предполагают, что ядерный ТБ принимает участие в структурировании ядра и гетерохрома- тинизации отдельных рибосомальных генов (Sjoberg et al., 2006). ТБ может быть фосфорилирован в 30 сайтах, при этом в норме на 1 моль этого белка приходится 2-3 моля фосфатных групп, тогда как у гиперфосфорилированной изоформы - от 5 до 9 молей фосфатов (Alonso et al., 2006; Kopke et al., 1993).Эти результаты убедительно демонстрируют ситуацию, когда сайты фосфорилирования присоединяют в норме по одному фосфату. В случае высокой интенсивности синтеза эти же сайты могут присоединять несколько фосфатов. Также показано, что N-гликозилирование характерно для гиперфосфорилированного ТБ, тогда как О-гликозилирование - для немодифицированного ТБ (Arnold et al., 1996; Wang et al., 1996). Предполагается, что фосфорилирование и О-гликозилирование этого белка играет роль в его ядерной локализации, а N-гликозилирование гиперфосфорилированного ТБ способствует транспорту этих комплексов к мембране нейрона.
12.
Еще по теме Тау-белки нейронов:
- Роль тау-белка в развитии БА
- Взаимосвязь между Ap и тау
- 15 Нейрон.обьединение нейронов
- Образование нейрофибриллярных сплетений из гиперфосфорилированных тау-белков
- Белки теплового шока и БА
- Белки
- Глава 5 Мембранные белки
- V- 3. 4. ВЛИЯНИЕ ЦР НА ФУНКЦИОНАЛЬНЫЕ БЕЛКИ
- Периферические мембранные белки
- Формирование пусковых факторов амилоидоза нейронов и активация теплозащитных механизмов латентной стадии болезни Альцгеймера
- Белки теплового шока
- Белки теплового шока 70
- Белки теплового шока в системе самозащиты почки