ВЫДЕЛЕНИЕ
Процессы выведения из организма конечных продуктов обмена веществ у иксодовых и аргасовых клещей, как и в других группах периодически питающихся кровососущих членистоногих, подчинены периодике гонотрофического ритма имаго и линочных циклов неполовозрелых фаз.
Кроме продуктов выделения, в ректальный пузырь, за исключением некоторых видов аргазид (Ornithodoros moubata), поступают продукты переваривания крови хозяина и разрушающиеся клетки средней кишки, а во время питания в значительном количестве мало измененная кровь. В результате фекалии клещей представляют смесь из нескольких веществ, соотношение между которыми меняется в разные периоды жизненного цикла.Состав экскретов. Конечный продукт азотистого обмена клещей — гуанин (Schulze, 1955; Kitaoka, 1961в), и в этом отношении они сходны с другими паукообразными (Schmidt a. oth, 1955). Гуанин отличается очень низкой растворимостью и выпадает в осадок уже при небольших концентрациях. В результате в мальпигиевых сосудах и ректальном пузыре он находится преимущественно в виде взвеси или кашеобразной массы кристаллов, на выведение которых из организма расходуется небольшое количество воды. В период эмбриогенеза, линьки или длительного голодания, когда клещи лишены возможности получать достаточное количество воды извне, плохая растворимость гуанина обеспечивает возможность его прогрессирующего накопления в мальпигиевых сосудах и предотвращает повышение его концентрации в гемолимфе до токсических значений.
Кристаллы гуанина имеют ярко-белую окраску и обладают интенсивным свечением в поляризованном свете. В содержимом мальпигиевых сосудов и ректального пузыря по внешнему виду можно различить мелкие (2—4 мк), не имеющие правильной формы, средние (10—20 мк) и крупные (40—80 мк) сфериты. Последние отличаются хорошо выраженной концентрической слоистостью и бывают простыми, двойными или сложными, т.
е. склеенными из нескольких простых (рис. 63). Кроме сферитов гуанина, в мальпигиевых сосудах питающихся особей довольно многочисленны шаровидные тела размером до 100 мк, образованные из более мелких эозинофильных шаров. Последние достигают диаметра в 1—3 мк и одновременно встречаются в цитоплазме клеток.Функционирование мальпигиевых сосудов. Биохимические нуги синтеза гуанина, как и место его образования в организме клещей, требуют дальнейших специальных исследований. В то же время прижизненные наблюдения над отпрепарованными мальпигиевыми сосудами и просмотр серийных срезов клещей Argas persicus, Ornithodoros papillipes (нимфы, самки и самцы), Hyalomma asiaticum и Ixodes ricinus (личинки, нимфы и самки) позволили выявить ритмику работы органов выделения.
Аргасовые клещи. У недавно перелинявших или длительно голодавших аргасовых клещей просвет мальпигиевых сосудов содержит большое количество сферитов гуанина, а клетки стенок умеренно уплощенные (рис. 335 стр. 193). После линьки происходит лишь частичная разгрузка сосудов от гуанина и в дальнейшем до питания вновь идет их постепенное заполнение экскретом. Сразу после питания наблюдается почти полное выведение гуанина из полости сосудов (фаза разгрузки; рис. 336). Одновременно увеличивается высота эпителиальных клеток стенок, вероятно, активно участвующих в выведении продуктов обмена, которые должны в большом количестве накапливаться по мере переваривания свежей порции белковой пищи. В течение нескольких дней после питания выделение гуанина в просвет сосудов не приводит к их заполнению сферитами благодаря быстрому вымыванию последних в ректальный пузырь и частым дефекациям. Позднее запас воды, полученный с кровью хозяина, исчерпывается, интенсивность дефекации ослабевает и просвет сосудов вновь постепенно заполняется гуанином (фаза загрузки) вплоть до следующего кровососания.
Иксодовые клещи. У только что перелинявших самок Hyalomma asiaticum и Ixodes ricinus мальпигиевы сосуды бывают заполнены большим количеством сферитов гуанина.
Разгрузка их от накопленных в период подготовки к линьке экскретов осуществляется в течение 1—3 дней после линьки. В дальнейшем на стадии послелиночного дораз- вития в просвете сосудов содержится небольшое количество одиночных мелких и средних сферитов, не образующих местных скоплений. Диаметр сосудов колеблется от 50 до 70 мк и они выглядят почти прозрачными.Эпителиальные клетки умеренной величины, кубические или слегка уплощенные (рис. 342).
У голодающих особей до прикрепления к хозяину наблюдается медленная загрузка полости сосудов сферитами гуанина. Последние образуют
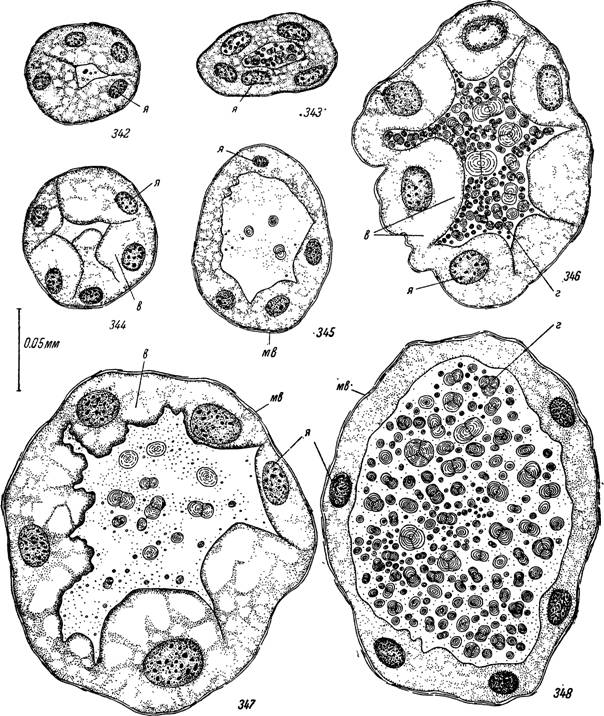
Рис. 342—348. Поперечные срезы мальпигиевых сосудов самки Ixodes ricinus на разных стадиях жизненного цикла.
342 — на стадии послелиночного доразвития; 343 — через 1 год голодания; 344 — на третьи сутки прикрепления, вес 10 мг; 345 — то же, участок, загруженный гуанином; 346 — напитавшаяся сразу после отпадения; 347 — перед началом яйцекладки; 348 — перед окончанием яйцекладки.
я — ядра эпителиальных клеток; мв — мышечные волокна; в — вакуоли; г — сфериты гуанина.
по ходу сосудов местные скопления (рис. 338), так что происходит чередование оптически пустых и белых (с гуанином) участков. Диаметр сосудов при этом существенно не изменяется. Клетки стенок сохраняют прежние размеры (рис. 343).
После прикрепления клещей к хозяину в первые 1—3 дня наблюдается освобождение сосудов от накопленных при голодании экскретов и они становятся полупрозрачными на всем своем протяжении (рис. 339). В то же время размеры эпителиальных клеток заметно увеличиваются и их апикальные концы местами вдаются в просвет (рис. 344—345). Диаметр сосудов увеличивается в 1.5—2 раза. Протоплазма в апикальной зоне вакуолизируется и местами в ней появляются эозинофильные включения. Размеры ядер заметно увеличиваются. Возобновляются митотические деления, но количество их меньше, чем при подготовке к линьке. Размеры клеток продолжают увеличиваться до конца питания и иногда по их апикальной границе выявляется палочковидная исчерченность.
Некоторые клетки подвергаются частичному разрушению (отторжение апикальных участков цитоплазмы) или даже полному.Постепенно, в связи с интенсификацией пищеварения, скорость отложения гуанина в мальпигиевых сосудах начинает превышать темпы его выведения в ректальный пузырь. Сфериты гуанина начинают вновь образовывать местные скопления (рис. 340). Ко времени окончания питания просвет сосудов уже на всем протяжении заполнен гуанином и органы приобретают характерную для них молочно-белую окраску. Стенки сосудов еще не подвергаются заметному растяжению, а сфериты гуанина свободно плавают в их жидком содержимом. Диаметр сосудов напитавшихся особей в 3—4 раза больше, чем у голодных (рис. 346). Подобный прирост достигается почти исключительно за счет роста и размножения эпителиальных клеток.
После отпадения с хозяина процесс загрузки сосудов гуанином продолжается с еще большей интенсивностью. Диаметр их на этой стадии может увеличиваться в 10 раз по сравнению с голодными особями. Они буквально на всем протяжении заполнены сплошной массой гуанина, который сильно растягивает их стенки (рис. 346—348). Ректальный пузырь на этой стадии также необычайно увеличен и забит одним гуанином.
У личинок и нимф процессы функционирования мальпигиевых сосудов протекают сходно с самками. Однако у них отсутствует столь сильное заполнение их гуанином благодаря периодическому освобождению от экскретов во время и после питания. При подготовке к линьке прямой кишки сообщение ректального пузыря с внешней средой прерывается. С этого момента и до окончания линьки дефекация отсутствует. Связь между мальпигиевыми сосудами и ректальным пузырем, напротив, не нарушается и в него непрерывно поступают большие количества гуанина. Размеры ректального пузыря к концу линьки необычайно увеличиваются и он занимает большую часть задней половины полости тела. Скапливающиеся в нем в огромном количестве сферокристаллы гуанина растягивают стенки до состояния мембрановидной оболочки с беспорядочно разбросанными уплощенными ядрами.
Растяжение стенок мальпигиевых сосудов и во время линьки, в отличие от напитавшихся самок, остается весьма незначительным (рис.
337). Перистальтические сокращения сосудов проталкивают скапливающийся в них гуанин в ректальный пузырь. Длина и диаметр сосудов значительно увеличиваются за счет делений и роста клеток их стенок (рис. 382). В результате число ядер, приходящихся на один поперечный срез через мальпигиев сосуд, увеличивается с 1—2 у личинок до 3—4 у нимф и5— 8 у самок.
У аргасовых клещей, по наблюдениям Л. К. Ефремовой (1967) над нимфами Alveonasus lahorensis, деления клеток мальпигиевых сосудов и рост органов наблюдаются на стадии линьки. Однако, в отличие от иксодид, последняя линька на имагинальную фазу не связана с делением клеток мальпигиевых сосудов. У взрослых аргазид размеры мальпигиевых сосудов уже не меняются и клеточные деления в их стенках отсутствуют. Увеличение размеров клеток у питающихся особей, возможно, связано с процессами их полиплоидизации. О полиплоидном характере ядер этих органов можно судить по появлению тетраплоидных наборов хромосом в делящихся клетках, но механизм этого процесса не исследован.
Ритмика дефекации. Освобождение ректального пузыря от скапливающихся в нем гуанина ж продуктов переваривания крови происходит с определенной цикличностью. У имаго аргасовых клещей наибольшее количество продуктов выделения выводится в первые дни после линьки и затем в течение 1—5 дней после кровососания. В то же время акты дефекации не прекращаются на протяжении всего гонотрофического цикла и сопровождаются выделением небольшой массы фекалий, состоящих, без особой закономерности, из гуанина (белая окраска), гема- тина или смеси обоих (черная окраска). Личинки и нимфы ведут себя сходным образом, но выделение фекалий у них постоянно прерывается на период от нескольких дней до нескольких недель перед линькой.
У имаго иксодовых клещей максимальные по объему количества гуанина выводятся в первые дни после линьки и во время питания, а у личинок и нимф и в первые несколько дней после его окончания. У самок после отпадения с хозяина дефекация сразу же прекращается и скапливающиеся экскреты остаются в организме до самой смерти клеща.
У напитавшихся личинок и нимф дефекация прерывается с началом отделения гиподермы от старой кутикулы.
Консистенция фекалий меняется в зависимости от содержания воды в организме. Во время питания или сразу после него они более жидкие, тогда как у голодных особей почти пылевидные. По-видимому, как и у некоторых других представителей членистоногих, клетки ректального пузыря способны к частичной реадсорбции воды.
Еще по теме ВЫДЕЛЕНИЕ:
- Выделение РНК (задача 1)
- Выделение фосфолипидов
- Выделение уридина
- 5 Выделение стрептококков
- Выделение цитидина
- Выделение бактериальной ДНК
- Обезвреживание фекалий и выделенных паразитов
- ПРАКТИЧЕСКОЕ ЗАНЯТИЕ № 2 Тема: Выделение ДНК из крови
- 4 Выделение золотистого стафилококка
- Выделение чистых культур
- 9.4. Выделение плазмидной ДНК.
- 8.2. Выделение продуктов ПЦР из агарозного геля.
- Выделение рекомбинантнного белка из биомассы
- ПРАКТИЧЕСКОЕ ЗАНЯТИЕ №3 Тема: Выделение ДНК на сорбент из крови
- Лечение вагинальных выделений, обусловленных раком шейки матки
- Выделение аденозина
- Микрофлора, выделенная у больных с гнойными воспалительными заболеваниями придатков матки.