Клітинна ланка імунітету
Такі фактори захисту організму від вірусних інфекцій, як віруснейтралізуючі антитіла, Р-інгібітори, температурна денатурація діють безпосередньо на вірус лише тоді, коли вірус, зруйнувавши клітини, виходить у міжклітинний простір.
Деякі віруси (наприклад віруси простого герпесу, цитомегаловірус) переходять із клітини в клітину по цитоплазматичних містках і, таким чином, уникають дії циркулюючих антитіл. У цьому випадку й у випадку інтегрованої форми інфекції, коли вірусний геном інтегрується в клітинну ДНК, основну роль у становленні імунітету відіграють клітинні механізми, пов'язані, у першу чергу, з дією специфічних цитотоксичних Т-лімфоцитів, Т-ефекторів ГЧУТ, НК і макрофагів (останні виявляють захисну дію й на ранніх етапах інфекції). Механізми клітинного противірусного імунітету в цілому дуже складні та базуються на засадах кооперації різних типів клітин - АПК, натуральних кілерів, Т-хелперів, цитотоксичних Т-лімфоцитів і гуморальних факторів: специфічних (антитіла) і неспецифічних медіаторів (ІНФ, цито- кіни, комплемент, інгібітори тощо). З клінічних спостережень відомо, що хворі з дефектами гуморального імунітету добре переносять вірусні інфекції, а особи з дефектами в Т-клітинній ланці імунітету страждають на вірусні захворювання, які є клінічним виявом цієї форми імунодефіциту.До основної популяції Т-клітин специфічної імунної відповіді належать ефектори цитотоксичних реакцій: Т-лімфоцити (CD8+, CD4+ цитотоксичні Т-клітини - ЦТЛ) і регуляторні CD4+ Т-хелпери типів І та ІІ.
CD8+ ЦТЛ експонують на поверхні клітини комплекс молекул: ТсИ (Т-клітинний антигенозв'язувальний рецептор) і корецептори CD3 та CD8. Саме ці молекули розпізнають вірусні антигени в асоціації з МНС-І на мембрані вірусінфікованих клітин. Важливу фізіологічну роль у регуляції диференціації ЦТЛ відіграє інтерлей- кін ІЛ-2, він також необхідний для індукції ЦДЛ.
ЦТЛ здійснюють прямий цитолітичний ефект з деструкцією клітини-мішені, окрім клітин імунопривілейованих органів (клітини гангліїв периферичної нервової системи, кришталика, тести- кул). Загибель клітин настає в результаті сил колоїдного осмосу.
Для лізису клітин необхідні такі умови:
• тісний контакт між ефекторною клітиною (ЦТЛ) і кліти- ною-мішенню;
• зв'язування ТсИ із антигеном;
• присутність двовалентних катіонів (магнію та кальцію), при цьому Mg2+ потрібен для зв'язування ЦТЛ із мішенню, а Ca2+ виконує подвійну роль, підсилюючи адгезію й забезпечуючи "летальний удар" - активацію перфорин-гранзимного шляху лізису клітини.
Перфорини та гранзими містяться в гранулах цитотоксичної клітини; перші утворюють отвори на мембранах клітин, другі (серинові протеази) руйнують білки (рис. 4.9, 4.10).
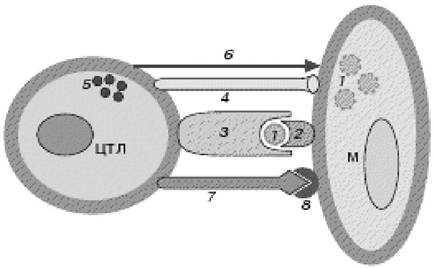
Рис. 4.9. Дія ЦТЛ на інфіковану клітину:
1 - вірусний антиген; 2 - МНС-1; 3 - антигенрозпізнавальний рецептор ЦТЛ;
4 - адгезійна молекула ЦТЛ; 5 - поляризація гранул ЦТЛ; 6 - секреція перфоринів і гранзимів ЦТЛ; 7-мембраноасоційована форма Fas;
8 - рецептор CD95 на клітині-мішені
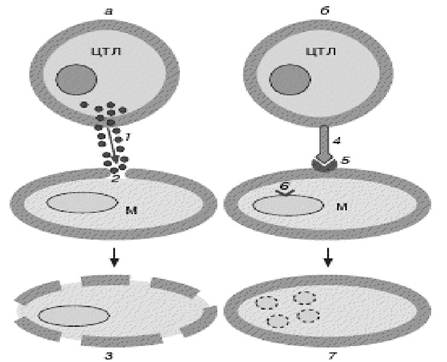
Рис. 4.10. Два шляхи загибелі клітини-мішені:
а) лізис (некроз); б) апоптоз.
1 - секреція перфоринів і гранзимів; 2 - утворення пор у мембрані клітини-мішені; 3 - загибель клітини шляхом лізису (некрозу);
4 - мембраноасоційована форма Fas; 5 - рецептор CD-95;
6 - індукція генів апоптозу; 7 - загибель клітини шляхом апоптозу
Процес лізису відбувається досить швидко (за 40-60 с). ЦТЛ стійкі до дії перфоринів і після взаємодії з клітиною-мішенню та її лізису можуть взаємодіяти з іншими мішенями. Цитотоксичні клітини використовують ще кілька ефекторних механізмів. Серед них індукція апоптозу - активація рецепторів загибелі клітин - Fas (CD95+).
Клітина гине протягом 4-6 год; апоптоз, опосередкований ФНП, ІНФ, перебігає повільніше й завершується через 18-24 год (рис. 4.11).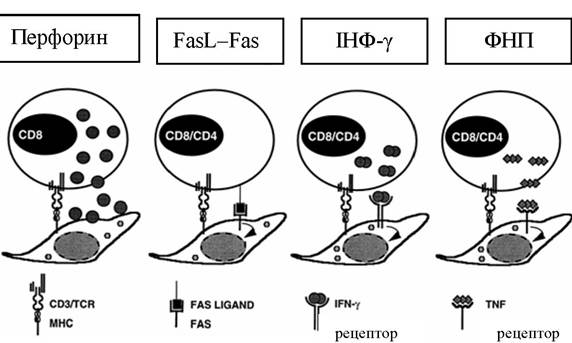
Рис. 4.11. Використання різних ефекторних механізмів цитотоксичними клітинами
Специфічні Т-кілери з'являються на ранніх термінах інфекції (через 1-3 доби), їх активність сягає максимуму через тиждень і повільно знижується протягом кількох тижнів, хоча клітини пам'яті цитотоксичних Т-лімфоцитів зберігаються тривалий час. Отже, утворення ЦТЛ є ранньою імунною відповіддю, яка часто передує утворенню циркулюючих антитіл.
Вірусні антигени як індуктори та мішені для ЦТЛ вивчені недостатньо. Однак відомо, що основною мішенню для ЦТЛ при респіраторно-синцитіальній інфекції є N-білок, тоді як G-білок не розпізнається цими клітинами. При вірусному гепатиті В провідну роль у розпізнаванні ЦЛТ відіграє білок серцевини, а при ЦМВ-інфекції - ранній структурний білок рр89. Індукторами ЦТЛ у вірусу простого герпесу є gpG, а у SV40 - великий Т-антиген. Установлено, що при грипозній інфекції в організмі інфікованого утворюються дві популяції ЦТЛ. Типоспецифічна популяція здатна розпізнавати й лізувати клітини, інфіковані різними штамами вірусу; штамоспецифічна розпізнає переважно гемаглютинін (НА) вірусу грипу. Типоспецифічний імунітет щодо споріднених до антигену вірусів грипу існує й за відсутності віруснейтралізуючих антитіл, що свідчить про важливу роль ЦТЛ у захисті від вірусів.
Імунітет, опосередкований ЦТЛ, виявляється по-різному при вірусних інфекціях. Вважають, що індукція вірусоспецифічних ЦТЛ при грипі, сказі - важливий компонент імунного захисту. ЦТЛ ефективно елімінують клітини, трансформовані вірусом Епштейна - Барр і вірусом хвороби Марека, ЦМВ, а також клітини, інфіковані вірусом вітряної віспи або гепатиту мишей. Т-лімфоцити, на думку багатьох дослідників, ефективніше пригнічують реплікацію вірусів, антигени яких експресуються на поверхні клітин на ранніх стадіях інфекції (віруси грипу, герпесу, альфа-віруси).
ЦТЛ менш ефективні в захисних реакціях проти тих інфекцій, збудники яких експресують свої антигени на пізніх (флавівірус) стадіях або зовсім не експресують їх (пікорна-, парвовіруси).Раннє та інтенсивне накопичення високоафінних і висо- коавідних вірусоспецифічних ЦТЛ - сприятлива ознака, що асоціюється з низьким вірусним навантаженням і здатністю імунної системи контролювати інфекційний процес; уповільнене - несприятлива ознака, що асоціюється з високим рівнем вірусного навантаження, хронізацією, прогресією хвороби та летальністю.
CD4+ Т-лімфоцити, окрім імунорегуляції всіх противірусних реакцій (опосередкованих як ЦТЛ, так і В-клітинами), можуть діяти як прямі ефекторні клітини (CD4+ЦТЛ), що експресують перфорин. Порушення в системі цитокінів, які контролюють напрям диференціації Т-лімфоцитів, призводять до руйнування кооперативних взаємодій імунокомпетентних клітин і порушень імунного гомеостазу. Унаслідок того, що протективна цитотоксична Т-клітинна відповідь викликає пошкодження в інфікованих клітинах хазяїна, рівновага між швидкістю розповсюдження вірусу та кінетикою Т-клітинної відповіді буде визначати або імунний захист, тобто виведення вірусу з організму, або ж імунопатологію.
4.7.
Еще по теме Клітинна ланка імунітету:
- Гуморальна ланка імунітету
- 3.1. Канцерогенез на рівні клітини
- Наступна це система імунітету, захисту.
- Взаємодія вірусів із клітинами
- Специфічний або набутий противірусний імунітет
- Молекулярні основи дії ІНФ на клітину
- Андрійчук О. М.. Вірусні інфекції людини та тварин: епідеміологія, патогенез, особливості противірусного імунітету, терапія та профілактика : навч. посіб. / О. М. Андрійчук, Г. В. Коротєєва, О. В. Молчанець, А. В. Харіна. - К. : Видавничо-поліграфічний центр "Київський університет',2014. - 415 с., 2014
- Вірусна інфекція та інтерферон
- ЗАПИТАННЯ ДЛЯ САМОКОНТРОЛЮ
- Шляхи виділення вірусів з організму
- СИНДРОМ БЕХЧЕТА
- Механізми та фактори персистенції вірусів
- 6.3.3. Імунотерапія
- ЛІМФОГРАНУЛЕМАТОЗ
- Роль факторів природної резистентності при вірусних інфекціях
- Функціональний атипізм.