Молекулярні основи дії ІНФ на клітину
ІНФ гальмує реплікацію РНК та ДНК-вмісних вірусів. Віруси, які чутливі до ІНФ, припиняють вірусоспецифічний синтез білків. Дія ІНФ на папілома-, поліомавіруси та деякі ретровіруси спостерігається на ранніх і пізніх стадіях репродукції.
ІНФ пригнічує первинну транскрипцію міксо- й рабдовірусів. Аденовіруси та вірус натуральної віспи стійкі до ІНФ, оскільки кодують білки-антагоністи, що блокують дію ІНФ.Протягом гострого інфекційного процесу активація ІНФ відбувається шляхом послідовних подій.
У процесі еволюції багатоклітинних організмів при розв'язанні задач розпізнавання й захисту від патогенів з'явились молекули-рецептори, здатні розпізнавати консервативні ознаки всіх класів мікроорганізмів. Ці рецептори називаються Toll-подібними. За допомогою 11 типів Toll-рецепторів клітини імунної системи, у першу чергу професійні фагоцити, упізнають консервативні компоненти практично всіх мікроорганізмів (у загальній формі полімери органічного походження чужорідні для багатоклітинного організму). Ліганд-рецепторна взаємодія запускає весь комплекс реакцій природного імунітету.
Природні імунні реакції зазвичай запускаються при розпізнаванні мікробних компонентів патогенозв'язаних молекулярних зразків (PAMPs), таких як ліпополісахариди (ЛПС) грамнегати- вних бактерій, 3-1,3-глікан грибів, пептидоглікани грампозитив- них бактерій і генетичні матеріали (ДНК або РНК) вірусів. Запуск ініціює два різні сигнальні шляхи. Перший опосередковується через Toll-подібний рецептор 3 (TLR-3). Ендосомна експресія TLR-3 розпізнає длРНК вірусного походження після потрапляння в клітину шляхом ендоцитозу. Другий шлях виконується цитозольними білками: протеїнкіназою R (PKR), RIG-1(retinoic acid-inducible gene) та MDA-5 (melanoma differentiation-associated gene 5). Ці імунні сенсори (RLR-RIG-like receptor) усередині клітин детектують вірусні геноми або реплікатив- ну діяльність вірусів.
RIG-I чутлива до 5'-трифосфатного мотиву вірусного геному або до коротких молекул длРНК, тоді як MDA-5 розпізнає довгі молекули длРНК (типовий проміжний компонент протягом реплікації деяких вірусів).
Хоча TLR-3- та RLR-шляхи розрізняються за ініціюючими стимулами, вони сходяться в точці активації транскрипції ядерних факторів, Nf-kB (nuclear factor kB), АР-1, G-білків, IRF3/7 (interferon regulating factor), які транслокуються в ядро та зв'язуються з чутливими до них послідовностями ДНК - генами ранньої відповіді. Стимуляція цих генів важлива для виходу клітини із Go-фази й переходу в G1-фазу, подальшої прогресії клітинного циклу, швидкого виробництва інтерферону типу I (ІНФ-ц та -β) та інших прозапальних цитокінів, які сприяють успішному розвитку адаптивного противірусного імунітету (рис. 4.3).
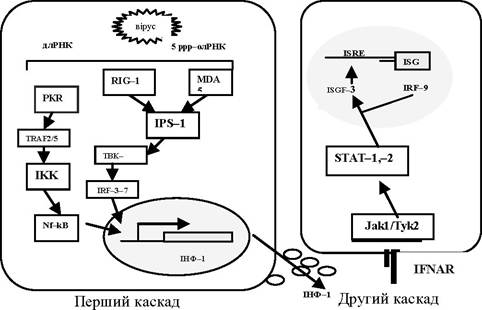
Рис. 4.3. Схема вірусної індукції системи ІНФ
Після завершення першого каскаду реакцій, який закінчується секрецією зрілого ІНФ, починається другий, пов'язаний з передаванням сигналу в сусідні неінфіковані клітини. ІНФ взаємодіє зі спеціалізованими рецепторами, які відрізняються для ІНФ типів І та ІІ (рис. 4.3). Після адсорбції та зв'язування молекул ІНФ на поверхні клітин через спеціалізовані рецептори (IFNAR) відбувається активація протеїнкіназ JAK-1, TYK-2. Вони фос- форилюють сигнальні трансдуктори транскрипції SТАТ1 та STAT2, які після цього взаємодіють із факторами стимуляції ІНФ IRF-9 (INF-regulated factor) і формують у ядрі гетеродимер- ний білковий комплекс ISGF3 (INF-stimulated gene factor). Цей комплекс зв'язується зі специфічними послідовностями ДНК, і в клітинах починають експресуватися до 30 генів ІНФ-індуцибельних білків, активуються латентні ферменти. Серед останніх у противірусному захисті беруть участь PKR, 2,5-олігоаденілатсинтетаза (2,5-ОАС), РНК-специфічна аде- нозиндезаміназа (АДАР) і гуанін-3-фосфатази (ГТФази, Мх- білки). Саме ці білки обумовлюють комплексний ефект клітинної резистентності до вірусів.
Центральною ланкою в антивірусному захисті клітин від вірусної інфекції є длРНК-залежна PKR, яка стимулюється длРНК і клітинним стресом. У клітині PKR локалізується переважно в цитоплазмі в асоціації з рибосомами. У латентному стані це мономер, афінний до длРНК. Активація PKR обумовлена безпосереднім зв'язуванням із длРНК, формуванням димерної структури, що викликає фосфорилування фактора ініціації трансляції еІ F 2 (це викликає його дезактивацію), оскільки ц-субодиниця еІ F 2 є специфічним субстратом для PKR. У результаті синтез білка в клітині припиняється. Це блокування однаково ефективне як щодо власних, так і щодо вірусоспецифі- чних мРНК. ІНФ типу І, з одного боку, індукує синтез PKR, а з іншого - активує цей фермент. PKR задіяна в противірусному захисті, регулює активність внутрішньоклітинної сигнальної трансдукції, інгібує проліферацію клітин.
Першою ознакою активної репродукції вірусу є внутрішньоклітинне накопичення длРНК (реплікативні форми геномної РНК). Одночасно длРНК є алостеричним активатором PKR. Утворення комплексу вірусоспецифічних длРНК із PKR фактично смертельно для клітини. У зв'язку з цим патогенному вірусу необхідні додаткові механізми забезпечення трансляції власних мРНК. Дійсно, такий механізм існує, його використовують як віруси грипу типу А різного ступеня патогенності, так і особливо небезпечні віруси Ебола, гепатиту С.
Другий шлях реалізації активності ІНФ - індукція синтезу 2,5-оліго-А-синтетази. Молекула 2,5-ОАС має длРНК-зв'язуючий домен і після взаємодії з вірусною РНК каталізує синтез моно-, ди-, тетраполіаденілатів. Ці продукти, у свою чергу, активують ендогенну латентну РНКазу L, яка селективно гідролізує вірусо- специфічні мРНК, тим самим забезпечуючи можливість трансляції клітинних мРНК.
Третій шлях пов'язаний з білком Мх - продуктом індукції ІНФ. Експресія Мх блокує вірусну реплікацію шляхом інтерференції з транскрипцією вірусів грипу, везикулярного стоматиту, кору та інших (-)РНК-геномних вірусів, його ефекту достатньо для пригнічення вірусної реплікації навіть за відсутності інших противірусних протеїнів.
Цей білок інгібує вірусну РНК- полімеразу, блокуючи синтез мРНК вірусу, а також інтерферує з транспортом вРНП-комплексу з цитоплазми в ядро.На рівні PKR вирішується доля інфікованої клітини: баланс вищеописаних процесів визначається на самому початку інфекції. Якщо рівновага буде переміщуватись у бік трансляції вірусних мРНК, то гіперактивна PKR разом із ІНФ індукуватимуть елімінацію інфікованої клітини шляхом апоптозу. Отже, виконується правило: якщо інфікована клітина не може бути санована від вірусу системою ІНФ, то вона повинна загинути. Елімінація інфекційного агента відбувається за рахунок ліквідації частини інфікованих клітин.
Інтерферони виконують важливі функції на першому рівні захисту від вірусної інфекції. При потраплянні вірусу до організму підвищення продукування ІФН є найшвидшою реакцією організму у відповідь на зараження інфекційним агентом. Система ІФН формує захисний бар'єр на шляху вірусів набагато раніше, ніж специфічні захисні реакції імунітету, за рахунок стимуляції резистентності клітин, роблячи їх непридатними для розмноження вірусів. Продукування ІФН-ц, ІФН-Р є першим етапом ранньої цитокінової реакції на інфікування вірусом. При високому вірусному навантаженні, зниженні резистентності організму, несприятливій дії факторів навколишнього середовища настає наступний етап цитокінової реакції, що характеризується активацією CD4+ та CD8+ Т-клітин із формуванням Т- і В-клітинної імунної відповіді та синтезом таких цитокінів, як інтерлейкіни ІЛ-1, ІЛ-6, ІЛ-10, ІЛ-15, ІЛ-18 і фактор некрозу пухлини. Останній підвищує продукування ІФН-у, який активує макрофаги, що мігрують до осередку вірусної інфекції та беруть участь у антивірусному захисті. ІФН-у є ключовою ланкою в неспецифічному та специфічному імунному захисті при вірусних інфекціях. Він чинить регулювальний вплив на більшість клітин імунної системи та їх взаємодію між собою, індукує антивірусну активність ІФН-а та ІФН-р. Таким чином, природний неспецифічний імунітет на клітинному рівні стримує реплікацію вірусів на ранній стадії, що дозволяє організму виграти час і в стислі терміни активувати адаптивний імунітет, необхідний для повного знищення інфекційного агента.
У цілому при проникненні вірусів у клітини розвиток інфекційного процесу прямо залежить від ефективності роботи системи ІНФ і здатності клітини-хазяїна протистояти репродуктивним і антиінтерфероновим механізмам вірусів.
На жаль, адаптаційні механізми вірусів випереджають еволюціонування імунної системи. Нині накопичена достатня інформація про складні взаємовідносини вірусів і системи інтерферонів. Ці відносини залежать від будови та особливостей репродукції вірусів, алгоритму індукції та дії основних типів ІНФ, здатності вірусів пригнічувати або уникати дії ІНФ, участі прозапальних і антизапальних цитокінів у антивірусному захисті. Усе це разом визначає патогенез певних вірусних інфекцій і результат захворювання.
Найдетальніше досліджені взаємовідносини системи ІНФ з вірусами грипу, гепатиту С та деякими іншими актуальними представниками царства Vira. Описані три групи вірусних інгібіторів системи ІНФ:
Інгібітори індукції (для першого каскаду) - це білки вірусів NS1, V (параміксо-), NS3-4 (ВгС), Є1(Ханта), E6 (папілома-), M (ВВС), v-IRF (ВГЛ-8).
Інгібітори дії (другий каскад) - C, V (параміксо-), P (сказ), VP24 (Ебола).
Інтерференція з продуктами ISG (інтерферончутливі гени) - короткі структуровані вРНК аденовірусів, ВГС, ВІЛ, ВЕБ; діють як псевдосубстрати, які інактивують клітинну PKR.
Наприклад, вірус грипу використовує два шляхи подолання впливу центральної ланки антивірусного захисту - активації PKR. Один шлях - можливість специфічного зв'язування вірусного NS-1-білка із длРНК (NS-1 належить до неструктурних білків, його синтез передує реплікації геномних РНК та мРНК, що і створює пул длРНК. Накопичення длРНК індукує протеїн- кіназу R). Вірусний білок інтерферує з PKR, при цьому не відбувається активація таких важливих факторів запуску генів ІНФ, як Nf-kB, АР-1, IRF3/7.
Інший шлях інактивації PKR - активація латентного білка р58 (належить до шаперонів), який перешкоджає димеризації PKR (фермент активний лише у вигляді димеру). Отже, цей білок є природним інгібітором PKR.
Вірус гепатиту С інгібує систему ІНФ, використовуючи такі механізми:
• Білок NS5А селективно пригнічує длРНК-залежну PKR у сайті димеризації. Крім того, білок блокує активацію фактора IRF, який регулює експресію генів ІНФ. Це найбільш загальний механізм пригнічення ІНФ серед РНК-геномних вірусів.
• Білок Е2 також пригнічує активність PKR, конкуруючи з її природним субстратом - фактором ініціації трансляції еІР 2α. Такий механізм використовує вірус віспи.
• Протеаза ВГС 3/4А інгібує активацію IRF-3 та пригнічує синтез ІНФ.
• При інфекції ВГС спостерігається блокування ІНФ-індукова- ного сигнального шляху JAK ^ТАТ.
Множинність механізмів інактивації противірусних ефектів ІНФ і продуктів його індукції в інфікованих клітинах свідчить про те, що ВГС високоадаптований до реакцій неспецифічних факторів захисту. В умовах різного рівня експресії вірусоспеци- фічних білків цей вірус може або пригнічувати систему клітинного захисту, або ж ухилятись від реакцій імунної відповіді. Основна боротьба відбувається між системою ІНФ і білками вірусу, що блокують дію ІНФ на різних рівнях. Слід зазначити, що ВГС порівняно з іншими вірусами становить значно більшу небезпеку щодо імуносупресії, оскільки PKR є для нього подвійною мішенню, а генетична детермінованість стійкості до ІНФ дублюється ще й геномом поверхневого антигену Е2.
Більшість РНК-вмісних вірусів кодують білки-інгібітори сигнальної системи ІНФ (табл. 4.2).
Таблиця 4.2
Приклади блокування чи обмеження продукування ІНФ вірусами
| Вплив | Вірус |
| Деградація длРНК | Вірусної діареї ВРХ (Erns має активність РНКази) |
| "Арешт" длРНК | Вісповакцини (E3), Рео (sigma3/A), Рота- (NSP3), грипу A (NS1), ВПГ (US11), ЦМВ (TRS1), ЦМВ мишей (m142∕14), ВЕБ (VP35), ВЕБ (SM) |
| Інгібування ТоІІ- рецепторної сигналізації | ВГС (NS3∕4a), вісповакцини (A52, A46) |
| Інгібування MDA-5 /RIG-I | Параміксо-^), грипу A (NS1) |
| Інтерференція з VAK | Хвороби Борна (P), вісповакцини (N1) |
| Інгібування деградації IRF-3 | РабдодТ). Хантаан (G1), ВПЛ-6 (E1), Рота-(NsP1), вірусної діареї ВРХ (Npro), класичної чуми свиней (Npro), ВПЛ-16 (E6), ВПГ (ICP0), ВГ ВРХ-1 (bICP0), РС ВРХ, РС, SARS Co-V |
| Інтерференція з IKK-комплексом | ВПЛ (E7), ВГС (core), вісповакцини, адеію-('Р3) |
| Інгібування NF-kB | Африканської чуми свиней (A238L), вісповакцини (N1), міксоми кролів (MNF) |
| Інтерференція з промотором активації IFN-b | ВГЛ-8 (IRF мімікрин) |
Ці білки здійснюють різні сценарії в сигнальному каскаді активації генів ІНФ, але основною мішенню для багатьох високопатогенних вірусів є протеїнкіназа R, що свідчить про її ключову роль у контролі внутрішньоклітинних сигналів.
Цікавими прикладами є особливо небезпечні вірусні інфекції. У цих вірусів ідентифіковано вірусоспецифічні антагоністи ІНФ. Наприклад, у вірусу денге виявлено три білки: NS2Λ, NS4Λ та NS4b, які пригнічують переважно передавання сигналу від рецепторів ІНФ типів І та П. У вірусу непальської гарячки два білки-антагоністи (V-білок пригнічує сигнальну систему активації, NS1 блокує продукування ІНФ-І та PKR), а вірус Ебола має унікальний білок VP35, який блокує вірусіндуковане фосфоритування фактора регуляції IRF-3 (експресії генів) ІНФ типу І.
Аналіз патогенезу інфекційних уражень, викликаний цими особливо небезпечними вірусами, показав, що, з одного боку, віруси мають численні механізми пригнічення ІНФ і деяких цитокінів, а з іншого - ці віруси спричиняють цитокіновий "вибух". Активний викид прозапальних цитокінів призводить до системного ураження внутрішніх органів і фактично разом з вірусами формує синдроми, що характерні для термінального стану, який часто закінчується смертю. Наприклад, віруси Ебола та Ласса інфікують альвеолярні макрофаги й дендритні клітини, які починають активно синтезувати прозапальні цитокіни (ІНФ, ФНП, ІЛ-2, ІЛ-10) у високих титрах. Підвищений рівень таких цитокінів викликає індукцію апоптозу як інфікованих, так і неінфікованих клітин. Крім того, ураження мононуклеарів і дендритних клітин не сприяє ефективній презентації антигену, необхідній для формування імунної відповіді. Отже, основним фактором патогенності, зокрема для зоонозних інфекцій, є генетично контрольована вірусним геномом здатність до пригнічення системи ІНФ.
Загалом противірусні ефекти інтерферонів зумовлені:
• універсальністю: дія на систему синтезу нуклеїнових кислот та білка;
• вираженою тканинною специфічністю;
• післядією: в оброблених ІФН клітинах тривалий час зберігається здатність пригнічувати розмноження вірусів;
• нечутливістю до індукованих вірусами антитіл.
4.4.
Еще по теме Молекулярні основи дії ІНФ на клітину:
- Клітинна ланка імунітету
- 3.1. Канцерогенез на рівні клітини
- Взаємодія вірусів із клітинами
- Мішені дії антивірусних агентів
- Дії у випадку загрози виникнення радіаційної небезпеки:
- Дії у випадку раптового виникнення радіаційної небезпеки:
- Назаренко Г. И., Осипов Г. С.. Основы теории медицинских технологических процессов. Ч. 2. Исследование медицинских технологических процессов на основе интеллектуального анализа данных. - М.: ФИЗМАТЛИТ,2006. - 144 с., 2006
- 15.1-15.2. Основы менеджмента
- Принципы в основе создания Изделия
- 1.2. Законодательно-правовые основы экспертизы трудоспособности
- Основы власти: