3.1. Канцерогенез на рівні клітини
Процеси, що призводять до трансформації нормальної клітини в ракову, є центральними в розумінні канцерогенезу. Рак є значною мірою клітинною проблемою. Без ракової клітини немає раку.
У ракову клітину може трансформуватися лише інтермітотична клітина, тобто така, що має камбіальний пул і зберегла здатність розмножуватися (епітелій, гермінативні і кровотворні тканини). Тканини, які не мають гермінативного пулу і не діляться (наприклад, центральної нервової системи), не можуть трансформуватися у злоякісні. Найчут- ливішою є клітина у фазі М, менш чутливою у фазах G1, G2 і нечутливою у фазі G0. Механізм канцерогенезу на клітинному рівні багатоступінчастий, тобто основні фази канцерогенезу (ініціація, промоція) мають ще «підфази», які залежать від якісних характеристик самих канцерогенів і від особливостей окремих клітин, зокрема фаз їх клітинного циклу.
Механізми хімічного та фізичного канцерогенезу як основних ініціаторів раку можна описати в спрощеній, схематизованій формі, вирізняючи тільки основні компоненти.
Вважається, що порогових (допустимих) концентрацій як хімічних, так і променевих канцерогенів немає і визначити їх неможливо. Причиною цього є наявність величезної кількості канцерогенів у навколишньому середовищі і потреба враховувати їх синергічну дію. Майже всі зовнішні хімічні канцерогени (первинні) вимагають ферментативної активації в клітинах, де вони перетворюються в кінцеві (ультимальні) канцерогени. Кінцеві канцерогени пошкоджують насамперед мембранні клітинні структури (зовнішню, лізосомну, епісомну мембрани, ретикулум, мітохондрії, апарат Гольджі) і остаточно геном клітини. Там вони зумовлюють різні пошкодження, починаючи від одного нуклеотиду (точкова мутація) і закінчуючи їх групами та генами (генні мутації). Активація первинних канцерогенів здійснюється ферментами, які виходять із
Рис.
3.1Схема хімічного канцерогенезу
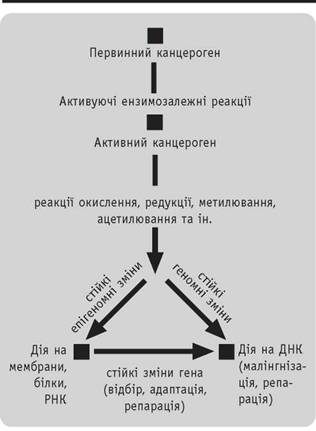
пошкоджених органоїдів клітини (переокислення, ацетилювання, метилювання (рис. 3.1). Наприклад, 3, 4-бензпірен-2-ацетиламінофлюорен перетворюється в його епоксидну форму— 2-ацетиламі- нофлюоренсульфат, вінілхлорид— у хлоретилен- оксид. Кінцеві канцерогени пошкоджують ДНК і РНК (мутагенна дія). Більшість таких пошкоджень підлягає репарації спеціальними ферментами. Ре- стриктази вирізають пошкоджені частини РНК і ДНК. РНК-полімерази синтезують комплементарні ділянки, а лігази зшивають новоутворені ДНК або РНК у суцільний ланцюжок. Таким чином, пошкоджений геном найчастіше відновлюється.
Аналогічний процес відбувається і при фізичному канцерогенезі. В його основі лежить радіоліз води, поява радикалів (H% HO^) з утворенням пероксиду водню, який взаємодіє з різними ферментами, різко порушуючи нормальний метаболізм клітини з вторинним пошкодженням генома. Крім цього, радіація безпосередньо може пошкоджувати ДНК і РНК, особливо а-променями і нейтронами, що теж призводить до мутації.
Вірусні білки (фосфопротеїди) спричинюють деструкцію клітинного каркасу (актин, вінкулін, фі- бропектин, мембранні білки), що сприяє розриву міжклітинних зв'язків. Ьольовані одна від одної клітини, щоб вижити, мобілізують гени, які продукують фактори росту епітелію, фібробластів, нервів, судин тощо.
Звичайно фактори росту отримують назви від тканин, де вперше були виявлені, хоча це має відносне значення для характеристики їх справжньої фізіологічної ролі. Наприклад, епітеліальні фактори росту є високоактивними при розмноженні як фібробластів, так і епідермальних та інших епітеліальних клітин. В одному клітинному середовищі фактор росту може бути потужним мітотичним фактором, в іншому— виконувати цілком іншу функцію. Характер дії факторів росту визначається не так ним самим, як місцем і умовами дії його на клітину в певний проміжок часу.
Фактори росту належать до найсильніших відомих біологічних субстанцій, що беруть участь у контролі фізіології клітин.
У багатьох клітинних культурах вони є активними в концентрації порядку 10...12 М і кількостях декількох пікограм (1 пі- кограм = 1 мільйонній частині мікрограма), викликаючи високоспецифічну відповідь. Будь-яка інтер-— 18 —
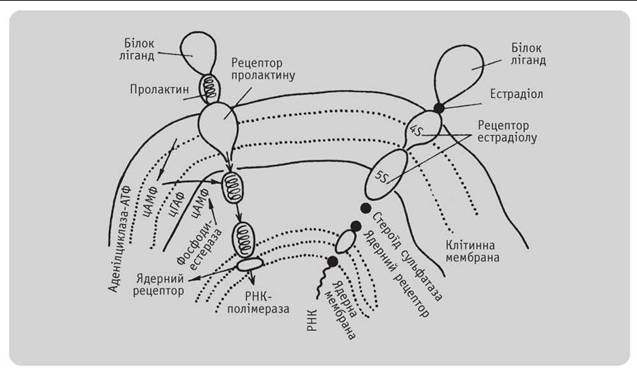
мітотична клітина реагує з факторами зовнішнього середовища за допомогою мембранних гліко- протеїдних рецепторів (рис. 3.2). Насамперед це стосується взаємодії між білковими факторами (гормони, фактори росту і трансформації, вірусні білки, протеолітичні ферменти). Рецептор реалізує програму. Багато з відомих рецепторів цього ряду мають тирозинкіназну активність, пов'язану з факторами росту. Фактори росту малі і дуже стійкі, їх рецептори значно більші, але лабільні. Тому відомості про рецептори факторів росту значно скупіші. Рецептори— складова частина мембрани. Дуже мало відомо про природу сигналу між рецепторами і внутрішнім середовищем клітини. Фактори росту— мультифункціональні, можуть діяти на багато клітин-мішеней. Вони можуть стимулювати клітинну проліферацію, гальмувати її при певних умовах або регулювати функцію, що не має відношення до проліферації.
Ракові клітини мають властивість автокринної секреції, тобто ракова клітина продукує ростові фактори, що реагують із власним відповідним рецептором. Якщо клітина сама продукує фактори росту і має до нього рецептор, вона позбувається зовнішнього контролю. У результаті ракові клітини потребують для максимального росту і поділу в культурі менше екзогенних факторів росту, ніж нормальні клітини.
Отже, фактори росту сприяють неконтрольо- ваному, не обмеженому в часі росту трансформованих клітин, тобто їх іморталізації. Подібну роль можуть відігравати гормони, які маюь рос- тостимулюючу дію (соматотропін, соматомедин, інсулін, естрадіол, гонадотропіни). Ростові фактори і гормони може виділяти трансформована клітина, незалежно від її гістогенетичної приналежності, тому що в кожній клітині є повноцінний набір усіх генів. Завдяки такій автокринній секреції може підтримуватися неконтрольований, автономний ріст, розмноження клітин з подальшим виникненням злоякісної пухлини.
Ріст трансформованої клітини може гальмуватися особливими білками (халонами), зокрема на ранніх стадіях розмноження ракових клітин.
Рис. 3.2
Схема Дії стероїдного (естраДіол) і білкового (пролактин) гормонів на клітину
— 19 —
Таким чином, виникнення ракової клітини не можна вважати якимось екстраординарним явищем у природі. Вся еволюція живого світу полягає в стійких мутаціях генів. Внаслідок мутації змінені клітини або гинуть (найчастіше), або пристосовуються до умов, даючи початок новим поколінням клітин і організмів. У раковотрансформованій клітині виникає фенотип, що характеризується ослабленням здатності клітини утворювати стійкі синцитіальні агрегати, можливістю жити і розмножуватися ізольовано, що породжує тенденцію до метастазування, і різними морфофункціональними рисами (понад 100), що описують ці властивості. До найсуттєвіших рис належать зміна співвідношення між об'ємом ядра і цитоплазми за рахунок збільшення ядра, його гіперхроматофілії, збільшення ядерець (тобто збільшення кількості ДНК і РНК) з появою нового генотипу. Серед епігеном- них змін слід назвати порушення співвідношення між гладким і шорстким ретикулумом, зниження стійкості мембранних структур, особливо зовнішньої мембрани (голі ядра), зміну ферментного складу клітин з порушенням їх функцій, як загальних (дихання, гліколіз), так і спеціальних, тобто продукування характерних для певної клітини специфічних ферментів і гормонів. Універсальною функцією ракової клітини є неповне спожиття нею глюкози з порушенням співвідношення між аеробним і анаеробним гліколізом, частіше на епігеномному і геномному рівні виникає новий тип клітин, які ми називаємо раковими. Їм властиві такі характеристики:
•Усі ракові клітини походять з нормальної клітини.
•У популяції ракових клітин більше таких, що перебувають у стадії мітозу, ніж у нормальній тканині, з якої вони походять.
•Ракові клітини, на відміну від нормальних, не здійснюють корисних для організму функцій.
•Ракові клітини живуть довше, ніж нормальні; культура ракової тканини в експлантаті характеризується необмеженим ростом, що дає змогу говорити про «безсмертя» (іморталізацію) ракової тканини. Отримано цікаві дані про роль апоптозу (програмованої загибелі клітин) у рості пухлин і появі низки біологічних особливостей. Феномен апоптозу в пухлинній культурі як правило гальмується.
• Ракові клітини не утворюють звичних тканинних структур, характерних для тканини, з якої вони вийшли, а схильні до хаосу.
•Ракові клітини, на відміну від нормальних, не залишаються на «власній території», а пенетрують у сусідні тканини.
• Ракові клітини мають тенденцію залишати своє початкове місце і поширюватися у віддалені ділянки організму.
Виникнення ракової клітини можливе лише на фоні нестабільності генома і недостатності регуляторних впливів, що контролюють проліферацію і диференціацію клітин. Нестабільність генома, як і гомеостазу організму,— це основний фон еволюції, який створює передумови для виникнення ракової клітини. Змінити цей закон нам не під силу, ми можемо лише сприяти тому, щоб звести до мінімуму фактори, що впливають на трансформацію нормальної клітини в ракову (хімічні та фізичні канцерогени). Серед людської популяції необхідно визначити групи ризику з генетичними та імунними недостатностями, у яких ракова трансформація буває частіше. Тому велике значення має пізнання канцерогенезу на рівні органа і організму.
Еще по теме 3.1. Канцерогенез на рівні клітини:
- 3.2. Канцерогенез на рівні органа
- 3.3. Канцерогенез на рівні організму
- Рівні управління та групи менеджерів
- Пропаганда, комунікація та соціальна мобілізація на місцевому рівні
- 46.Етапи та рівні соціалізації особистості в у різних теоретичних концепціях.
- Стадии канцерогенеза.
- 2.6. Воспаление и канцерогенез
- 5.1. Полимерный канцерогенез: некоторые исходныефакты и положения
- Канцерогенез
- 5.3. Канцерогенез, индуцируемый природными иискусственными минеральными частицами
- КАНЦЕРОГЕНЕЗ КАК КОМПЛЕКСНОЕ ИЗМЕНЕНИЕ ГЕНОТИПА КЛЕТКИ
- Глава 14 КАНЦЕРОГЕНЕЗ
- Мутационный канцерогенез.
- Розділ 3. Канцерогенез
- Молекулярно генетические аспекты канцерогенеза при раке желудка
- Роль метилирования в канцерогенезе
- Роль вирусов в канцерогенезе.
- Современные представления о канцерогенезе и патогенетических механизмах развития рака гортани.