Гуморальна ланка імунітету
Гуморальна відповідь - утворення антитіл (рис. 4.5) - розвивається переважно на поверхневі (структурні) і деякі внутрішні (неструктурні) білки вірусів. Імуногенність вірусних білків залежить від їх хімічної структури, молекулярної маси та численних факторів хазяїна.
Інфікуюча доза вірусу, переважний шлях введення та, відповідно, потрапляння в імунну систему вірусних антигенів суттєво впливають на кінетику та спектр індукованих антитіл.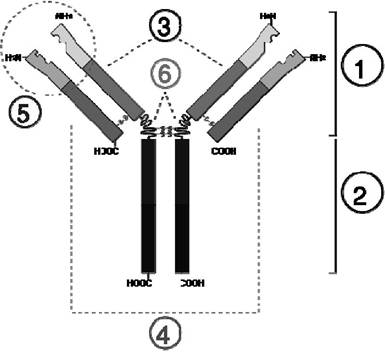
Рис. 4.5. Структура IgG: 1) Fab (fragment antigen binding - антигенозв'язувальний фрагмент); 2) Fc (fragment crystallizable фрагмент, здатний до кристалізації); 3) важкий ланцюг; 4) легкий ланцюг;
5) антигенозв'язувальна ділянка; 6) шарнірна ділянка
В експериментальних дослідженнях установлено, що внутрішньовенне введення вірусів грипу стимулює більш виражену імунну відповідь, ніж інтраперітонеальне або підшкірне. У процесі антитілогенезу залучаються багато типів клітин - дендритні клітини, В- та Т-лімфоцити. Реакція В-лімфоцитів починається зі зв'язування вірусних (чужорідних) білків з молекулами поверхневих рецепторів (імуноглобулінів), що експресуються на мембрані наївних В-клітин. Певний тип В-клітин генетично запрограмований на експресію рецепторів тільки однієї специфічності, тобто розпізнає тільки один епітоп, локалізований у молекулі антигену. Інтенсивність утворення сироваткових і секреторних антитіл перебуває під контролем Т-системи лімфоцитів. CD4+ Т-лімфоцити, що мають на мембрані ТсЯ тієї самої специфічності, що й антигеноспецифічні В-клітини, розпізнають даний комплекс, активуються та у відповідь продукують цитокіни - ІЛ-2, ІЛ-4-ІЛ-6, ІЛ-10, а також гамма-інтерферон (рис. 4.6).
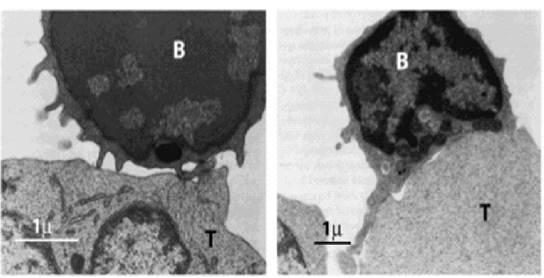
Рис. 4.6.
Два шляхи активації B-клітин:ліворуч - тимусонезалежна активація антигеном; відповідає за антитілоопосередковану або гуморальну імунну відповідь; праворуч - тимусозалежна активація антигеном і прямий контакт із Т-хелперами. Антигени, які активують цей шлях, називають тимусозалежними
Кількість, спектр і співвідношення цитокінів детермінують інтенсивність проліферації та диференціації активованих В- лімфоцитів у плазматичні клітини - продуценти антитіл. Плазматичні клітини у результаті стимуляції синтезують значну кількість противірусних поліклональних антитіл. Специфічність та афінність (ступінь специфічної спорідненості активного центра антитіл до антигенної детермінанти) антитіл під час імунної відповіді суттєво зростають. Одночасно з накопиченням плазматичних клітин і пропорціонально їх кількості утворюються епітопоспецифічні В-клітини пам'яті. Аналогічні закономірності спостерігаються й при формуванні імунної відповіді на імунізацію.
Водночас при деяких вірусних інфекціях у біологічних рідинах у гостру фазу антитіла ледве визначаються. Потім упродовж двох - трьох тижнів після закінчення інфекції їх біосинтез різко зростає, що відповідає вторинній імунній відповіді, побічно підтверджує перенесену інфекцію й визначає резистентність до повторного зараження. Антитіла з противірусною активністю можуть бути виявлені в різних рідинах організму - плазмі або сироватці крові, носовому, глотковому, бронхіальному секретах, цереброспінальній і слізній рідинах, спермі, вагінальному секреті, випорожненнях. Розподіл класів антитіл у них варіює. У сироватці крові зазвичай визначаються всі три класи антитіл - IgG (домінують), IgM, IgA. Більшість із них синтезована плазматичними клітинами кісткового мозку, лімфатичних вузлів і селезінки. У рідинах слизових переважають антитіла IgA, вони синтезуються плазматичними клітинами лімфоїдної тканини, що асоціюється зі слизовими оболонками. Визначаються також антитіла IgG, частина яких є результатом транссудації білків із кровоносного русла, а частина синтезована локально.
Динаміка гуморального імунітету виявляється, у першу чергу, у зміні синтезу антитіл різного класу (рис. 4.8). Перші етапи імунної відповіді на інфекцію відповідають інкубаційному періоду. На початку продромального періоду в інфікованих осіб синтезуються вірусоспецифічні антитіла IgM - основа первинної імунної відповіді. Продукування антитіл починається доволі швидко, за короткий час (один - два тижні) їх рівень досягає високих значень і у два - вісім разів перевищує рівень антитіл IgG-класу. мають відносно слабку спорідненість до антиге
ну. Наприкінці двох - трьох тижнів уміст IgM- та IgG-антитіл у сироватці крові практично однаковий. Рівень антитіл IgM деякий час ще залишається таким самим, потім протягом 20-30 днів різко знижується, а через 1-3 місяці після завершення інфекції ці антитіла можуть повністю зникнути. Однак якщо титр IgM після 14 днів незмінний або підвищений, то це свідчить про продовження інфекції.
Поствакцинальні IgM зникають із кровотоку повільніше, ніж постінфекційні. Слід зазначити, що розвиток первинної гуморальної імунної відповіді (продукування антитіл) досягає піка повільніше, ніж накопичення антигеноспецифічних ефекторних Т-клітин.
^М циркулюють переважно в плазмі крові, лімфі; здатні до преципітації, аглютинації та лізису антигенів (рис. 4.7). Вони активно зв'язуються з комплементом, не проходять крізь плаценту, тому виявлення IgM-антитіл у плоді свідчить про внутрішньоутробну інфекцію (червона висипка, цитомегалові- русна інфекція, парвовірусна хвороба свиней тощо). Існування цих антитіл у крові нетривале, період напіврозпаду становить вісім - десять днів.
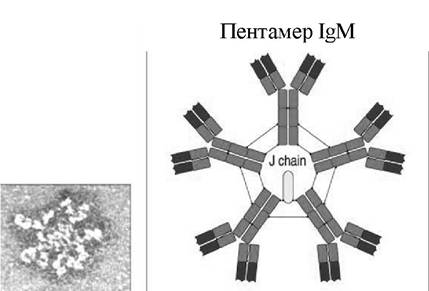
Рис. 4.7. Структура IgM
Численні серологічні маркери одного й того самого вірусу дають змогу слідкувати за перебігом інфекційного процесу, а визначення циркулюючих антитіл до різних компонентів вірусу дозволяє розрізнити гостру інфекцію (табл. 4.5), стан персисту- ючого носійства та період одужання.
Таблиця 4.5
Визначення форми ВЕБ-інфекції за серологічними маркерами
| Форма інфекції | IgM VCA | IgG VCA | IgG EAD | IgG EBNA |
| Гостра стадія первинної інфекції | + | + | + | - |
| Нещодавно перенесена інфекція | - | + | + | + |
| Давно перенесена інфекція | — | + | — | + |
| Хронічна інфекція | +/- | + | + | +/- |
| Реактивація | +/- | + | + | +/- |
У період розквіту захворювання відбуваються як якісні, так і кількісні зміни імунної відповіді - переключення антитілоутво- рення плазматичними клітинами з IgM на IgG, інтенсивніше їх продукування, дозрівання й підвищення афінітету. Синтез IgG настає через три - сім днів після зараження або імунізації. Продукування IgG має триваліший характер порівняно з ІдМ, особливо після природного інфікування або імунізації живою вакциною (віспа, поліомієліт, чума великої рогатої худоби).
Стійкі високі титри антитіл до певних структур вірусу відображають також персистенцію вірусу (кір, ВЕБ) або повторну стимуляцію антигеном, але антитіла до деяких вірусів можуть тривалий час циркулювати й без чітких ознак захворювання.
Вміст IgG у сироватці крові під час хвороби найвищий (12 г/л); із сироватки крові вони можуть потрапляти в тканинну рідину. IgG долають плацентарний бар'єр, забезпечуючи плід материнськими антитілами, які зникають у новонародженої дитини протягом перших 12-15 місяців життя.
IgG формують імунологічну пам'ять.Концентрація IgG інтенсивно зростає протягом двох - чотирьох тижнів, сягає піка через півтора - три та більше місяців і певний час підтримується на постійному рівні. Надалі вірусо- специфічні видо- та серотипоспецифічні IgG-антитіла залишаються основним ізотипом антитіл, які здійснюють контроль інфекції в організмі.
Період напіврозпаду IgG становить 25-35 днів. Однак якщо титри IgG зберігаються на високому рівні довше, то це може свідчити, що інфекція триває. Для повного одужання кількість антитіл має перевищувати вміст віріонів і вірусних білків. У таких випадках вірусне навантаження швидко знижується, зникають клінічні симптоми інфекції.
Природне інфікування респіраторними й кишковими вірусами супроводжується утворенням секреторних і сироваткових IgA.
Продукування специфічних антитіл IgA починається дещо згодом, темпи їх накопичення повільніші, а досягнення піка (не дуже високого) припадає на четвертий - шостий місяць. IgA представлені у двох формах: сироваткові IgA - у вигляді мономеру, тоді як секреторні IgA є димером (рис. 4.8). Мономе- рна молекула синтезується плазматичними клітинами кісткового мозку, периферичних лімфовузлів, селезінки та інших органів. Димерні форми синтезуються лімфоїдною тканиною шлунково- кишкового або респіраторного трактів. При цьому сироваткові антитіла цього типу становлять > 70, а секреторні - гепатиту А - клітинні ліпіди тощо). Необхідно пам'ятати, що імунна відповідь не лише сприяє одужанню й забезпечує захист від повторного зараження вірусом, але й може викликати патологічні зміни в організмі хворого. Наприклад, у присутності антитіл до вірусу денге, які не мають нейтралізуючої активності, вірус розмножується швидше, і таке імунне посилення реплікації є причиною більш тяжкого перебігу хвороби. Є повідомлення про те, що при незначній концентрації нейтралізуючих антитіл інфекційність деяких вірусів посилювалась щодо клітин, що мають на поверхні Fc-рецептор. Імунопатологічні реакції спостерігали й при імунізації проти кору та РС-інфекції.
Природне інфікування на тлі недостатності імунітету внаслідок застосування слабкоанти- генних інактивованих вакцин супроводжувалось тяжчим перебігом захворювання, ніж у людей, що не були вакциновані.Дія антитіл, окрім нейтралізації вірусу (табл. 4.7), полягає в тому, що вони викликають руйнування інфікованих вірусом клітин (феномен імунного лізису). Можливі два варіанти такої цитотоксичності. Комплементозалежна цитотоксичність виникає при дії антитіл на антигени, що експресовані на поверхні інфікованої клітини, з подальшою активацією системи комплементу. Відбувається збірка комплексу, що лізує мембрану клітини. Комплементозалежний цитоліз можливий лише при високій щільності експресії вірусних антигенів на клітинній мембрані (5 х 106 на клітину). На противагу цьому для лізису за механізмом АЗЦК необхідна присутність на клітині-мішені лише 103 молекул IgG - така кількість забезпечує зв'язування НК.
Оцінка гуморальної імунної відповіді. Серологічні дослідження з метою виявлення противірусних антитіл у крові, оцінка динаміки їх утворення та рівня (титру) широко застосовується в лабораторній практиці. Головними типами досліджуваних антитіл є нейтралізуючі, інгібуючі гемаглютинацію, комплементофік- суючі, імунофлуоресциюючі, преципітуючі, цитотоксичні. Для оцінки антитілоутворення частіше використовують варіанти імуно-ферментного аналізу як найбільш специфічного, чутливого, кількісного, швидкого та автоматизованого. Усе ширше в діагностиці та диференціальній діагностиці застосовується метод виявлення антитіл за допомогою імуноблоттингу, а також метод імунохроматографічних стрипів тощо.
Уникнення вірусом нейтралізуючої дії антитіл у популяції хазяїна. Віруси, що викликають гострі летальні інфекції, не здатні уникнути нейтралізуючої відповіді антитіл або інших ефекторних механізмів імунної системи, оскільки біологічний вид хазяїна і сам вірус припиняють існування. Унаслідок цього такі віруси на короткий час беруться імунною системою під контроль, вірусам необхідно найшвидше передати інфекцію неімунному організму (стратегія "ударити та втекти"). Оскільки кількість чутливих неімунних хазяїв у популяції зменшується при кожному передаванні, то гострі цитопатогенні віруси повинні мати спосіб ухилитися від уже існуючого популяційного імунітету. Це досягається шляхом інфікування інших видів і розширення спектра антигенних і протективних детермінант. При цьому віруси утворюють серотипи, або підтипи (хоча різні серотипи мають багато спільних детермінант, нейтралізуючі епітопи в них різні). Епідеміологічні спостереження у випадку інфекцій, викликаних вірусом поліомієліту або грипу, указують на те, що серотипоспецифічний імунітет не дає перехресного захисту, хоча вірус везикулярного стоматиту та поліовірус мають стабільні серотипи. Навпаки, вірус грипу типу А динамічно змінюється для того, щоб уникнути дії популяційного імунітету. Завдяки постійним мутаціям поверхневих білків (антигенний дрейф) або повній їх заміні шляхом реасортації відповідних генів (антигенний зсув - шифт) вірус грипу типу А удосконалив здатність ухилятись від дії популяційного імунітету. Антигенний шифт значно збільшує випадки захворюваності вірусом грипу (розвиток пандемії) і рівень смертності. Варіанти вірусу, що виникли внаслідок дрейфу, здатні підсилювати відповідь антитіл пам'яті на попередній вірус, яким був праймований організм, хоча вже існуючі нейтралізуючі антитіла не захищають від нового варіанта вірусу.
Уникнення вірусом дії антитіл в організмі хазяїна. Схильність до персистенції дозволяє вірусам зменшувати цикли передавання, необхідні для їхнього виживання, що дає їм змогу уникнути повного контролю імунної системи. Такі віруси створили власні механізми уникнення гуморальної імунної відповіді - мутації віруснейтралізуючих детермінант під час інфекції. Отже, саме імунна відповідь організму зумовлює постійну селекцію нових варіантів вірусу, які не елімінуються, в організмі окремого хазяїна.
Узагальнюючи, можна сказати, що віруси (такі як вірус грипу) уникають імунної відповіді в популяції хазяїна шляхом антигенного дрейфу, а віруси, подібні до ЛХМ та ВІЛ, використовують механізми антигенної варіації в організмі окремого хазяїна. Унаслідок цього з'являються численні квазівиди, що не схожі на вірус, яким первинно був інфікований організм.
Однак слід зазначити, що антитіла не здатні впливати на розвиток і розмноження вірусів у клітині. Антитіла малоефективні також при захисті організму від хронічних і повільних інфекцій. Це вказує на те, що стимуляція антитілоутворення при більшості вірусних інфекцій є невиправданою процедурою та не приводить до суттєвого підвищення противірусного імунітету. У цілому вона використовується лише для запобігання реінфекції.
Формування інфекційних імунних комплексів (ІК). При взаємодії вірусів з антитілами можуть утворюватись імунні комплекси, у яких віруси зберігають інфекційну активність. Розрізняють два типи ІК:
1) Утворюється в організмі на ранніх етапах інфекційного процесу, коли в крові присутня незначна кількість противірусних антитіл, але з часом кількість антитіл зростає, і комплекси знищуються. ІК із залишковою інфекційністю можуть формуватися при гальмуванні антитілоутворення або при імунодефіцит- них станах, які характеризуються пониженим рівнем синтезу імуноглобулінів.
2) Виявляють навіть при надлишку антитіл, рівень інфекцій- ності такого ІК становить 0,0001-10 % від початкової. Ця залишкова інфекційність отримала назву персистуючої (ненейтралі- зуючої) фракції. Саме такі ІК можуть дисоціювати, їх розглядають як один із факторів розвитку хронічних форм вірусного захворювання.
Тривала циркуляція в організмі таких комплексів призводить, по-перше, до постійного інфікування чутливих клітин, по-друге, - до антигенної стимуляції вірусоспецифічних імунокомпетент- них клітин. У результаті формуються нові ІК, які містять інфекційний вірус. Описано феномен посилення репродукції вірусів при використанні недостатньої концентрації антитіл для альфа-, флаві-, бунья-, рабдо- та реовірусів. Особливо чітко цей феномен виявляється при зараженні вірусом денге макрофагів, де зв'язаний антитілами вірус розмножується набагато краще, ніж вільний.
Імунологічна пам'ять Після інфікування або вакцинації хазяїна імунна система праймується. Така специфічна імунологічна пам'ять призводить до посиленого захисту проти повторної інфекції. Імунологічну пам'ять зазвичай оцінюють за підвищеними титрами антитіл або збільшеною частотою виявлення специфічних Т-клітин.
Отже, антитіла мають широкий спектр противірусної активності:
1) нейтралізують інфекційність віріонів;
2) блокують прикріплення віріонів до рецепторів клітин і проникнення (депротеїнізацію, піноцитоз);
3) опсонізація поверхні віріонів полегшує захват їх фагоцитами; підсилюють фагоцитарну реакцію;
4) взаємодіють з антигенами вірусів, що циркулюють у крові, утворюючи імунні комплекси;
5) активують систему комплементу, лізують оболонкові віруси та інфіковані клітини;
6) індукують АЗКЦ інфікованих вірусом клітин НК-клітинами, моноцитами та макрофагами.
4.6.
Еще по теме Гуморальна ланка імунітету:
- Клітинна ланка імунітету
- Наступна це система імунітету, захисту.
- Специфічний або набутий противірусний імунітет
- Показатели гуморального иммунитета
- Показатели гуморального иммунитетау обследованных женщин
- 35. Теории сна (сосудистая, гуморальная, теория центризма)
- Гуморальная и клеточная иммунные системы.
- Изучение состояния местного гуморального иммунитета у часто болеющих детей с патологией ЛОР-органов.
- Влияние на воспаление нервных и гуморальных факторов
- Методы изучения клеточного и гуморального иммунитета
- Клеточный и гуморальный иммунитет при послойной кератопластике
- Изучение состояния системного и антиген-специфического гуморального иммунитета у часто болеющих детей с патологией ЛОР- органов.
- Изучение местного гуморального иммунитета аденоидных вегетаций и небных миндалин у часто болеющих детей
- 5.3. Характеристика показателей гуморального иммунитета у больных урогенитальным хламидиозом
- Гуморальные агенты синдрома системного воспалительного ответа и их действие