Взаємодія вірусів із клітинами
Взаємодія клітин і вірусів - складний багатогранний процес, обумовлений багатьма різними факторами, зокрема типом і видом клітини та вірусу, ступенем чутливості (або резистентності) клітини, кількістю вірусних часток і чистотою популяції вірусу, що інокулюється, дією різноманітних фізичних і хімічних факторів, імунологічним і генетичним статусом макроорганізму та ін.
Типи взаємодії вірусів і клітин. Реакція клітин на вірусну інфекцію досить різноманітна. Запропоновано багато способів класифікації реакції клітин на зараження вірусами. Найбільш застосовною є класифікація вірусних інфекцій на клітинному рівні, що запропонована В. М. Ждановим та А. Г. Букринською (1986), відповідно до якої вірусні інфекції розподіляються на автономні та інтеграційні (рис. 3.1).
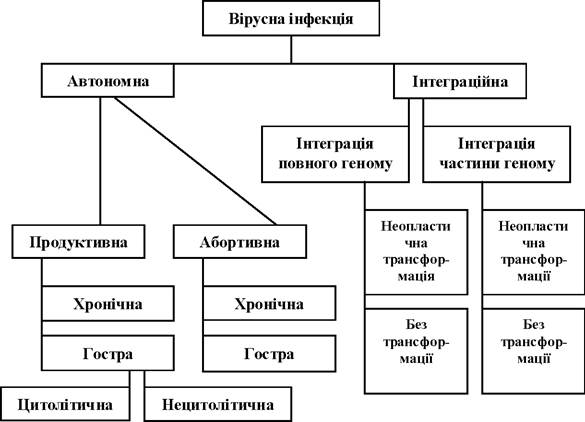
Рис. 3.1. Класифікація вірусних інфекцій на рівні клітини
Автономна - тип вірусної інфекції, при якій вірусний геном реплікується незалежно від клітинного. Поняття автономії відносне, воно обмежується лише відсутністю фізичного контакту між вірусним і клітинним геномами, хоча їх взаємодія постійно відбувається в процесі інфекції. Цей тип взаємодії характерний для більшості вірусів тварин.
Інтеграційна - тип вірусної інфекції, при якій вірусний геном частково або повністю інтегрується з клітинним геномом і реплікується разом із ним. Інтеграційна інфекція виникає в результаті фізичного об'єднання геному вірусу та клітини. При цій формі інфекції геном вірусу реплікується й функціонує як складова частина клітинного геному. Інтеграція може зумовити неопластичну трансформацію. Наприклад, трансформація поліомавірусів та їх онкогенні властивості є результатом експресії вірусоспецифічних ранніх генів і взаємодії з продуктами специфічних клітинних генів (p53, pRB). У трансформованих клітинах і клітинах пухлин ДНК поліомавірусів інтегрована в геном клітини.
Папіломавірусна ДНК часто, але не завжди, присутня в інтегрованому стані, переважно в цервікальних карциномах, у той час як для карциноми шкіри характерна наявність епісомної форми. Онкогенні типи вірусів папіломи людини (ВПЛ) надають клітинам можливості необмеженого поділу. Це є результатом дії генів E6 та E7. Е6 приєднується до р53 і призводить до його деградації, а Е7 взаємодіє з клітинним білком pRB та іншими спорідненими білками. Взаємодія вірусних онкобілків із клітинними інгібіторами циклінзалежної кінази (р16, р21, р27) - також важлива подія в процесі іморталізації.
Продуктивна вірусна інфекція завершується утворенням інфекційного потомства, на відміну від абортивної, для якої не характерне утворення інфекційних часток, або вони утворюються в значно меншій кількості, ніж при продуктивній вірусній інфекції. Абортивна інфекція розвивається при проникненні збудника в нечутливі клітини (наприклад, при проникненні вірусу лейкозу корів у організм людини) або в клітини, не здатні забезпечити повний репродуктивний цикл (наприклад ті, що перебувають у стадії клітинного циклу G0). Здатність клітин підтримувати вірусоспецифічні репродуктивні процеси також пригнічує ІФН.
Умови виникнення абортивної інфекції. 1. Зараження чутливих клітин дефектним вірусом. Дефектним називається вірус, не здатний виявити всі генетичні функції, які необхідні для утворення інфекційного потомства. Слід однак розрізняти терміни дефектні віруси та дефектні вірусні частки. Дефектні віруси - це віруси, що репродукуються лише в присутності вірусу-помічника. Наприклад, вірусом-помічником для аденоасоційованого вірусу (парвовірусу) є адено- або герпесвірус, для вірусу гепатиту Д - вірус гепатиту В. Дефектні вірусні частки накопичуються в популяціях багатьох вірусів, особливо при пасивуванні їх із високою множинністю інфекції. Дефектні вірусні частки інтерферують при репродукції вірусу з інфекційними вірусними частками, і тому отримали назву дефектні інтерферуючі частки (Ді-частки, або ДІЧ).
2. Зараження чутливих клітин у непермісивних умовах. В організмі непермісивними є такі умови: підвищення температури, зміна рН у вогнищі запалення та концентрація іонів, наявність антиметаболітів, інгібіторів тощо; в експерименті - зміна температури інкубації, складу поживного середовища, внесення антиметаболітів та інгібіторів тощо. У результаті або вірус гине без утворення інфекційних нащадків, або переривається на певному рівні. При зміні непермісивних умов на пермісивні абортивна інфекція перетворюється на продуктивну. Заміна абортивної інфекції на продуктивну може відбутися й за допомогою вірусу-помічника.
3. Зараження нечутливих клітин стандартним вірусом. Це найбільш розповсюджена форма абортивної вірусної інфекції.
4. Гостра вірусна інфекція - це така форма інфекції, при якій після утворення вірусного потомства клітина або гине, або видужує та не містить вірусних компонентів. При хронічній вірусній інфекції клітини продовжують продукувати вірусні частки або їх компоненти протягом тривалого часу й передають цю здатність спадково. Частіше хронічної форми набуває абортивна інфекція, оскільки вірусний генетичний матеріал зазвичай не входить до складу вірусного потомства, а накопичується в клітинах і передається дочірнім клітинам. Одним із факторів, що викликають хронічну інфекцію, є ДІЧ. Такі частки, потрапляючи в клітину разом з інфекційними вірусними частками, конкурують із ними за фактори продукування й перешкоджають утворенню інфекційного вірусного потомства. У результаті загибель клітин не відбувається. При появі в системі нових чутливих клітин у них знову виникає продуктивна вірусна інфекція з утворенням ДІЧ, і цикл повторюється.
Гостра інфекція, яка завершується загибеллю клітини (лізисом), називається цитолітичною, а гостра вірусна інфекція, яка безпосередньо не призводить до лізису клітини і при якій клітина ще може функціонувати впродовж деякого часу, продукуючи вірусні частки, називається нецитолітичною. Віруси, що викликають загибель клітини, називають цитопатогенними, а ураження клітини - цитопатичним ефектом.
Характер цитопатичних змін при вірусних інфекціях може бути різноманітним: дегенерація клітин (вакуалізація цитоплазми, пікноз ядер, хромосомні аберації), утворення внутрішньоклітинних тілець-включень, виникнення гігантських клітин (полікаріоцити, симпласти). Лізис клітини - кінцева стадія дії цитопатогенних вірусів. Для вияву цитопатогенності вірусу не потрібні відтворення геному та реалізація всієї генетичної інформації.
Нелітична автономна інфекція характерна для вірусів, що містять зовнішню ліпопротеїнову оболонку та вивільняються з клітини шляхом брунькування. Ретро-, ортоміксо-, параміксо-, аренавіруси здатні викликати продуктивну нелітичну інфекцію клітин. Проте в найтиповішому випадку нелітичний характер властивий абортивним інфекціям, коли відсутні або не виявляються генетичні властивості вірусів, необхідні для здійснення цитопатогенних функцій.
Змішана інфекція. У природних умовах розповсюджений феномен змішаної інфекції, при якій клітина заражується двома або кількома різними вірусами. Можливі варіанти взаємодії вірусів у процесі змішаної інфекції:
1. Один з вірусів пригнічує репродукцію іншого або пригнічується репродукція обох вірусів (інтерференція вірусів).
2. Вірус підсилює репродукцію іншого вірусу в результаті комплементації. Комплементація може відбуватися між двома спорідненими або неспорідненими вірусами, наприклад між аденовірусом та аденоасоційованим вірусом людини.
3. Обидва віруси суттєво не впливають на процес репродукції кожного, проте може відбуватися порушення морфогенезу обох вірусів.
Форми взаємодії вірусів і клітин. При взаємодії вірусу з клітиною можна виділити такі основні типи: цитопатичну, вірусопатичну, алобіофоричну, резистентну та вірусоцитопатичну взаємодії.
Цитопатична взаємодія завжди закінчується деструкцією клітини. У процесі взаємодії може утворюватися й тимчасово існувати новий біологічний комплекс вірус-клітини. У цьому комплексні вірусний геном може функціонувати більш або менш автономно, що виявляється переважно в тому, що репродукція вірусного геному здійснюється незалежно від геному клітини.
Зазвичай у цьому випадку спостерігається продуктивна вірусна інфекція. Однак іноді цикл розмноження може перериватися на будь-якій стадії онтогенезу вірусу, і повноцінне потомство не утворюється (абортивна інфекція).При цитопатичній формі спостерігається зазвичай літична інфекція. Деструкція клітин відбувається в деяких випадках після однієї лише адсорбції віріонів або навіть окремих компонентів на поверхні клітини (цитотоксична дія вірусу).
При цитопатичній формі взаємодії геном вірусу зазвичай не інтегрує з геномом інфікованої клітини.
У багатьох випадках зараження клітини вірусом супроводжується швидким (часто вибірковим) пригніченням синтезу клітинних макромолекул - ДНК, РНК, білків. На пізніших стадіях інфекційного процесу відбувається повне порушення метаболізму клітини, включаючи припинення дихання, гліколізу, а також будь-яких синтетичних реакцій. Наприклад, інфіковані вірусом клітини втрачають здатність до мітотичного поділу. Поряд з біохімічними та функціональними порушеннями в зараженій вірусом клітині розвиваються морфологічні зміни, які виявляються за допомогою методів світлочутливої мікроскопії (цитопатична дія (ЦПД)).
Віруси можуть викликати розвиток у клітині деструктивних процесів двома шляхами.
1. Цитопатична дія ззовні. Цитотоксичну дію можуть викликати позаклітинні віріони чи навіть окремі їх компоненти. При цьому навіть не обов'язково, щоб вірус потрапив усередину клітини. Наприклад, в аденовірусів білок пентон викликає агрегацію й відділення від скла клітин у культурі, а білки, що входять до складу гексонів і фібрил, різко знижують синтез ДНК, РНК та білків як у зараженій, так і в незараженій клітині.
2. Цитопатична дія зсередини. Деструкція клітин прямо чи опосередковано пов'язана з проникненням і функціонуванням у ній геному вірусу. Зміни метаболізму клітини, що розвиваються внаслідок цього, можуть бути результатом дії одного чи кількох факторів: пригнічення синтезу ДНК, РНК та білків. У процесі вірусної інфекції відбувається зміна субмікроскопічної організації клітини - гіпертрофія та деструкція клітинних органоїдів, формування нових внутрішньоклітинних структур та ін.
Деякі з цих структурних пошкоджень клітини викликаються безпосередньо вірусоспецифі- чними білками. Наприклад, структурні білки ентеровірусів беруть участь у розвитку пізніх цитоморфологічних змін.Причини цитопатичної дії (ЦПД) вірусів можна поділи на такі:
• порушення нормальної життєдіяльності клітин у результаті механічного пошкодження клітинних структур вірусними компонентами (дефекти цитоплазматичної мембрани, які виникають у результаті проникнення чи виходу вірусів із клітини);
• руйнування лізосом і вихід їх ферментів у цитоплазму (автоліз клітин);
• виснаження білкових та енергетичних ресурсів клітин за рахунок переключення клітинних ферментів і білоксинтезуючого апарату на синтез вірусоспецифічних компонентів;
• специфічне ушкодження вірусом клітинних молекул.
За терміном виникнення ЦПД поділяють на ранню та пізню. Рання ЦПД виявляється в перші години після інфікування (від 3 год після інфікування клітинних культур вірусом), спричиняється дією структурних елементів вірусів. У цей період ще не відбулося проникнення вірусу в клітину. Даний тип ЦПД морфологічно виявляється в порушенні клітинного моношару, заокругленні клітин, відокремленні їх від скла. Пізня ЦПД виявляється, коли вірус (чи його геном) потрапляє в клітину й зазвичай на кінцевих стадіях репродукції вірусів у клітині. Пізня ЦПД виявляється в утворенні полікаріоцитів і включень. Вирішальну роль у розвитку деструктивних процесів у клітині на пізніх стадіях можуть відгравати також клітинні білки, у тому числі гідролази лізосом. У результаті пошкодження лізосомальної мембрани в цитоплазму виходять різноманітні клітинні ферменти (ДНКаза, РНКаза, фосфатаза, протеаза та ін.), що й призводить до швидкого та повного руйнування інфікованої клітини.
Для характеристики деструктивних змін одношарових культур клітин, що заражені різними вірусами, найчастіше використовується робоча класифікація ЦПД (О. Г. Анджапарідзе, 1962):
Перша група. Рівномірно розподілена дрібнозерниста деструкція клітин. Виникає при ураженні клітин вірусами поліомієліту, Коксакі, ЕСНО, натуральної віспи, грипу тощо.
Друга група. Вогнищева дрібнозерниста деструкція клітин з тяжами незмінених клітин між ними. Виникає при ураженні клітин вірусами весняно-літнього енцефаліту, вірусом хвороби Ауескі (псевдосказу), спумавірусами тощо.
Третя група. Осередкові скупчення заокруглених клітин, що нагадують грона винограду. Виникає при ураженні клітин аденовірусами.
Четверта група. Рівномірно розподілена крупнозерниста деструкція клітин (клітини збільшені в розмірі, заокруглені). Виникає при ураженні клітин вірусом простого герпесу, вірусом В та ін.
П'ята група. Утворення гігантських багатоядерних клітин - симпластів і синцитіїв. Виникає при ураженні клітин вірусами кору, вісповакцини, герпесу та ін. При цьому типі ЦПД відбувається розчинення клітинних оболонок, унаслідок чого цитоплазма сусідніх клітин зливається, утворюючи єдине ціле, у якому (переважно по периферії) розташовані ядра клітин.
Слід розрізняти терміни симпласт і синцитій. Різниця між цими поняттями полягає в тому, що синцитії виникають унаслідок часткового злиття цитоплазматичних мембран клітин, на відміну від повного злиття мембран при утворенні симпластів. Таким чином, синцитій - це група клітин, об'єднаних протоплазматичними відростками, а симпласти мають загальну масу протоплазми, у якій міститься багато ядер.
При багатьох вірусних інфекціях у клітинах (у ядрі чи цитоплазмі) різних органів і тканин з'являються особливі утворення, які називають тільцями-включеннями. Їх класифікують за локалізацією в клітині, вмістом нуклеїнової кислоти, тинкторіальними властивостями та гомогенністю (гомогенні чи зернисті).
Тільця-включення локалізуються вибірково. Наприклад, цитоплазматичні включення характерні при віспі (тільця Гварні- єрі), сказі (тільця Бабеша - Негрі), грипі, парагрипі, чумі великої рогатої худоби тощо. При ринотрахеїті великої рогатої худоби, ларинготрахеїті птахів, аденовірусній інфекції, герпесвірусній інфекції (тільця Каудрі), ящурі розвиваються включення в ядрі. За природою включення можуть бути місцями утворення вірусних часток (фабрики віріонів - тільця Гварнієрі), скупченням вірусів (тільця Бабеша - Негрі, поліедри), глибками хроматину (тільця Каудрі).
Особливою формою деструкції клітин є імунний цитолітич- ний ефект (імунний цитоліз), що розвивається в інфікованих вірусом клітинах під впливом специфічних противірусних антитіл та дії ефекторних лімфоцитів. Існують дві форми імунного цитолізу:
• клітинний імунний цитоліз. Клітини-мішені, що містять антигени, індуковані вірусом, лізуються під впливом ефектор- них Т-лімфоцитів;
• гуморальний імунний цитоліз. Під впливом специфічних антитіл і комплементу розвивається швидке (кілька хвилин) ураження структур клітини.
Вважають, що імунний цитоліз є одним з основних механізмів, що забезпечують швидку деструкцію (у певних умовах) інфікованих клітин у зараженому організмі та нейтралізацію вірусу. При цьому не спостерігаються пікноз і фагоцитоз клітин, а відбувається їх розпад, що призводить до поширення в організмі вірусних і змінених клітинних антигенів. Останнє може сприяти розвитку аутоімунних та імунопатологічних захворювань.
Вірусопатична взаємодія вірусу та клітини характеризується збереженням і розмноженням клітин та деструкцією вірусу. Клітина резистентна до вірусу чи на стадії його адсорбції на її поверхні (наприклад у результаті інактивуючої дії клітинних рецепторів), чи на стадії проникнення в неї вірусу. В останньому випадку онтогенетичний цикл розвитку вірусу переривається через деструкцію вірусу, що проникнув усередину клітини, у гіпертрофованому фагосомно-лізосомальному апараті клітини. У цьому апараті можуть руйнуватися не лише віруси, що тільки потрапили в клітину, але й наново синтезовані віріони.
Алобіофорична форма взаємодії (носійство іншого життя) - це форма взаємодії вірусу й клітини, яка характеризується тривалим (протягом необмеженої кількості пасажів) співіснуванням геномів вірусу та клітини, що забезпечує збереження й розмноження цих видів живих істот і фрагментів їх геномів у природі. При цьому один вид - клітина - є носієм іншого виду - неклітинної форми життя (вірусу) або частини його геному, і навпаки, вірус може зберігати у складі свого геному фрагмент геному клітини.
Розрізняють два типи алобіофоричної форми взаємодії.
1. Інтегративна: інтеграція повного геному чи ДНК- транскрипту; інтеграція фрагмента геному чи фрагмента ДНК- транскрипту; інтеграція з продукуванням повного вірусу; інтеграція з утворенням неповного вірусу чи окремих вірусоспецифі- чних антигенів ; інтеграція з продукуванням нових клітинних антигенів; інтеграція без продукування повного вірусу та синтез вірусних чи клітинних антигенів; інтеграція без зміни метаболізму клітини; інтеграція з трансформацією клітини; включення в геном вірусу фрагмента (фрагментів) генетичного (хромосомного чи позахромо- сомного) апарату клітини.
2. Персистентна: персистенція з продукуванням повного вірусу; персистенція з продукуванням нових клітинних антигенів; персистенція без зміни метаболізму клітини; персистенція зі зміною метаболізму клітини.
При взаємодії вірусів із клітинами спостерігаються різні патологічні зміни, які зумовлені низкою факторів:
• особливістю структури вірусів;
• швидкістю репродукції вірусу та кількістю інфекційних часток у потомстві;
• чутливістю та пермісивністю клітин до вірусів;
• умовами культивування клітин (склад поживного середовища, рН середовища, температура культивування); на рівні організму - це його реакція на зміни клітин і тканин, які були викликані інфекцією.
Патологічні зміни уражених вірусами клітин зумовлені специфічними й неспецифічними процесами. До неспецифічних відносять процеси, обумовлені змінами деяких ланок обміну речовин, проникності цитоплазматичної мембрани; маргинацію хроматину, вакуолізацію цитоплазми, пікноз ядер, хромосомні аберації, а саме:
• дезинтегративне набрякання ядер клітин - феномен збільшення розмірів ядра клітини в результаті стимуляції обміну речовин при вірусній інфекції;
• цитотоксичну дію вірусів - вплив вірусів на клітину ще до стимуляції.
Усі ці зміни відносять до неспецифічної взаємодії вірусів із клітинами, тому що вони не залежать від дози вірусу, розмірів віріонів, типу клітин, характеру утворення включень, тривалості вірусної інфекції.
Специфічні процеси взаємодії вірусу з клітиною включають:
• різноманітні зміни, що виявляються в пригніченні синтетичних процесів, порушенні функціональної активності, пошкодженні структур клітини та її загибелі (ЦПД, утворення включень);
• цитопроліферативний ефект, який зазвичай призводить до трансформації клітин;
• резистентність клітин до вірусів;
• вірусопатичну дію;
• синтез неповноцінних віріонів і вірусних специфічних антигенів (за абортивної інфекції);
• індуктивний вплив (синтез інтерферонів).
Рівні взаємодії вірусів і клітин. Виділяють п'ять основних рівнів взаємодії клітин і вірусів: внутрішньоклітинний (молекулярний), клітинний, популяційно-клітинний, взаємодія на рівні багатоклітинного організму та на рівні популяцій макроорганізмів.
Молекулярний (внутрішньоклітинний). На цьому рівні внутрішньоклітинний вірус взаємодіє зі структурними елементами клітини. Ці взаємини залежать від локалізації вірусу.
Чужорідна для клітини генетична інформація вірусів залежно від їх природи може перебувати у вільному стані або локалізуватися у фаголізосомному апараті, у цитоплазмі клітини у формі плазміди чи в інтегрованому вигляді з хромосомним або поза- хромосомним генетичним апаратом клітини. В останньому випадку геном одного чи кількох ДНК-вмісних вірусів (чи ДНК- транскриптів) може інтегруватися в одну чи кілька ділянок однієї чи кількох хромосом клітини, а також у позахромосомний генетичний апарат клітини (ДНК мітохондрій і центріолей). У виникаючому при такій взаємодії комплексі вірус-клітина відбувається одночасна репродукція двох інтегрованих геномів, що підкоряється загальній регуляції. Наприклад, при дослідженні лімфобластоїдних клітин Raji було встановлено, що вони містять близько 50-60 геномів ВЕБ на клітину та існують у вигляді цитоплазматичних плазмід. Плазмідна ДНК перебуває під клітинним контролем (синхронна реплікація, відсутність більшості вірусоспецифічних функцій).
Клітинний. Визначається властивостями вірусу та клітини. У пермісивних для репродукції вірусу системах вірус адсорбується та взаємодіє з однією інтерфазною клітиною чи клітиною, що ділиться, одно- чи багатоядерною клітиною, симпластом чи гетерокаріоном (внутрішньо- чи міжвидовим клітинним гібридом).
Популяційно-клітинний. Визначається взаємодією вірусу з популяцією диплоїдних, клональних чи гетерологічних клітин in vitro, популяцією гетерологічних клітин в органній культурі чи органі багатоклітинного організму.
При таких формах взаємодії можливе пасивування та формування багатьох поколінь вірусів. Основною особливістю вірусної інфекції в клітинній популяції є гетерогенність системи у зв'язку з гетерогенністю вірусних часток і клітин, що входять до складу популяції. Гетерогенність виявляється за наявністю в популяції вірусу зрілих, неповноцінних, вегетативних форм, окремих компонентів вірусу, асоціацією вірусів із різних родин. Клітини в кожній клітинній популяції широко варіюють за чутливістю до вірусу; інфекція може перебігати не так, як на клітинному рівні. Наприклад, при ураженні вірусом, що викликає в клітині продуктивну інфекцію, чутливі клітини популяції можуть загинути, і в популяції за рахунок деякої кількості нечутливих клітин може встановитися хронічна інфекція.
Рівень багатоклітинного організму. Репродукція вірусу перебуває під контролем імунологічних механізмів гуморального та клітинного імунітету макроорганізму. Значну роль у формуванні резистентності організму до вірусу відграють генетичні фактори - спадковість.
Рівень популяції макроорганізмів. Визначається взаємодією популяції вірусів з популяцією ссавців, птахів, комах тощо. Значна роль у формуванні вірусної патології людей належить зоонозам, змінам оточуючого середовища та соціальним умовам життя.
Отже, по мірі переходу від найпростішого (внутрішньоклітинного) рівня до найскладнішого (взаємодії вірусів з популяцією макроорганізмів) значно збільшується кількість форм, варіантів і наслідків взаємодії вірусу з хазяїном. Крім того, проблема взаємодії популяцій вірусів і клітин може ускладнюватися за рахунок гетерогенності популяції вірусів і наявності в ній зрілих, неповноцінних форм вірусів та окремих їх компонентів, а також асоціацією неспоріднених вірусів (коінфекція, яка може призводити до інтерференції, посиленої репродукції обох вірусів).
3.4.
Еще по теме Взаємодія вірусів із клітинами:
- Тропізм вірусів
- 3.1. Канцерогенез на рівні клітини
- Клітинна ланка імунітету
- Механізми та фактори персистенції вірусів
- Поширення вірусів у організмі
- Молекулярні основи дії ІНФ на клітину
- Шляхи виділення вірусів з організму
- Збірні групи вірусів, що викликають масові інфекції
- РОЗДІЛ 1. Загальна характеристика родин вірусів людини та тварин
- Класифікація та загальна характеристика вірусних інфекцій на рівні організму
- ЗАПИТАННЯ ДЛЯ САМОКОНТРОЛЮ
- Вірусна інфекція та інтерферон
- Загальне уявлення про патогенез вірусних інфекцій
- ЗАПИТАННЯ ДЛЯ САМОКОНТРОЛЮ