Специфічний або набутий противірусний імунітет
Набутий противірусний імунітет формується в результаті реагування імунної системи на інфекцію, обумовлену генетичним або серологічним варіантом вірусу певного виду, і визначається унікальними властивостями імунних Т- та В-лімфоцитів вибірково відповідати на чужорідні антигени та формувати специфічну імунну пам'ять, яка виражається в посиленій і прискореній реакції на повторну зустріч із цим інфекційним агентом.
Віруси та їх антигени здатні викликати потужні неспецифічні реакції, стимулюючи набутий імунітет, або, навпаки, - імуносу- пресію та імунопатологію. Недостатність імунної системи хворого (найчастіше за дефіциту системи Т-клітин) і антигенна мінливість збудника часто супроводжується його персистенцією та хронізацією інфекційного процесу. Кожній інфекції притаманний власний спектр гуморальних і клітинних факторів захисту залежно від біологічних особливостей вірусу та його антигенного складу. Противірусний імунітет спрямований передусім на нейтралізацію та елімінацію з організму вірусу, його антигенів та інфікованих вірусом клітин.
Визначення специфічності, пам'яті та толерантності, ключові параметри імунної відповіді. Специфічний гуморальний імунітет опосередковується антитілами, які продукуються плазматичними клітинами й потрапляють у кров. Такі антитіла розпізнають конформаційні детермінанти в білках, вуглеводах, антигенах (таких як віруси та бактерії), на слизових оболонках та в крові. Попри все, за винятком місця пошкодження, антитіла не можуть проникати в щільні тканини. Клітинний імунітет опосередковується лімфоцитами, які утворюються зі стовбурової поліпотентної ЄВ34+-клітини в кістковому мозку, проходять низку етапів диференціації та дозрівання. Потім ЄВ3+Т- лімфоцити мігрують у тимус для завершення диференціації й дозрівання. У тимусі вони дивергують на дві субпопуляції зрілих, але наївних типів Т-клітин СВ3+СВ4+ (хелпери) і СВ3+ЄВ8+-лімфоцити (цитотоксичні).
Далі вони мігрують у периферичні лімфоїдні органи - селезінку, лімфатичні вузли, шкіру та підслизові лімфоїдні утворення. В-лімфоцити (CD19+, ЄВ20+-клітини) з кісткового мозку мігрують безпосередньо в кров і периферичні лімфоїдні органи.Розвиток імунної відповіді на вірусну інфекцію. Противірусний імунітет починає формуватись на стадії презентації вірусного антигену. Відповідь організму на збудник відбувається в чотири стадії (табл. 4.3).
Таблиця 4.3
Стадії розвитку імунної відповіді
| Стадії | Клітини, що залучені в розвиток процесу | Імунологічні процеси |
| Індукція | Макрофаги, дендритні клітини, клітини Лангерганса, антиген- реактивні лімфоцити | Процесинг і презентація антигену |
| Імунорегуля- торна (проліферативна) | Т-хелпери, Т-супресори, В-супресори | Активація та взаємодія іму- норегуляторних клітин. Проліферація та диференціація клітин |
| Ефекторна (продуктивна) | Т-кілери, Т-ефектори, плазматичні клітини | Накопичення та активація ефекторних клітин |
| Імунологічна пам'ять | Т- та В-клітини пам'яті | Накопичення клітин пам'яті |
Перша стадія індукції включає момент потрапляння антигену (вірусу) в організм, процесинг і презентацію його Т- та В -лімфоцитам.
Вірус, як і будь-який інфекційний збудник, є складним антигенним комплексом, до складу якого входять численні антигенні компоненти, кожний з яких викликає власний імунітет. Загалом імунна відповідь розвивається не на вірус або його білок, а на окремі пептиди, низькомолекулярні епітопи збудника. Захоплення вірусу та презентація його антигенів відбувається за участю професійних антигенпрезентуючих клітин (АПК) - макрофагів, дендритних клітин (ДК), клітин Лангерганса.
Ці клітини захоплюють циркулюючий у крові вірус або зруйновану вірусом клітину та мігрують до регіональних лімфовузлів, де й відбувається презентація чужорідного антигену Т- та В-клітинам.На всіх етапах Т-клітинної імунної відповіді, починаючи з антигенного розпізнавання, її індукції (активація Т-хелперів з активаційним маркером CD4+) і закінчуючи ефекторною ланкою (активація цитотоксичних Т-лімфоцитів - ЦТЛ з активаційним маркером CD8+), ключову роль відіграють молекули головного комплексу гістосумісності - загальна назва МНС (major histocompatibility comlex), у людини - HLA (human leukocyte antigens), у мишей - Н2 (histocompatibility).
Існує три класи молекул MHC: HLA класу I (HLA-A, HLA-B та HLA-C, HLA-G), які експресуються на більшості ядерних клітин, HLA класу II (HLA-D, HLA-DP, HLA-DQ, HLA-DR), які експресуються на антигенпрезентуючих клітинах (В-ліфоцитах, активованих макрофагах, дендритних клітинах). Продукти генів HLA класу III - це білки системи комплементу й деяких цитокінів.
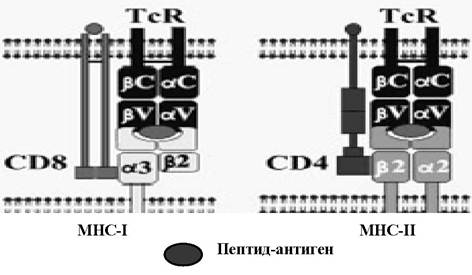
Рис. 4.4. Розпізнавання антигену (МНС -рестрикція)
У середині 80-х рр. було з'ясовано, що перед представленням Т-клітинам антиген проходить етап внутрішньоклітинної обробки за участю молекул МНС, які не лише презентують антиген, але й зв'язують його після гідролізу, а потім транспортують на клітинну мембрану. Цей процес називають МНС-рестрикцією, він забезпечує розподіл зусиль в імунній системі: Т-клітини спрямовані проти власних змінених клітин, що експресують МНС- молекули й чужорідні пептиди (рис. 4.4), а молекули антитіл, що продукуються В-лімфоцитами, елімінують позаклітинний антиген.
Існують два шляхи презентації антигену та способів його обробки. Це представлення ендогенного та екзогенного антигену. Ендогенний антиген - це антиген, який синтезувався в інфікованій клітині. Новоутворені вірусні пептиди піддаються деградації убіквітинзалежним шляхом у протеосомах до фрагментів довжиною біля дев'яти амінокислотних залишків. Далі ці фрагменти транспортуються в ендоплазматичний ретикулум білками-транспортерами TAP-1,2 (transporter associated with processing), де вони утворюють комплекс із молекулами МНС класу І.
Цей комплекс транспортується в апарат Гольджі, де відбувається глікозилування (стабілізація) ланцюгів МНС. Потім комплекс вбудовується в плазматичну мембрану для взаємодії з Т-клітинним рецептором (IcR) CD8+7- лімфоцитів (рис. 4.6). У тих випадках, коли клітина не інфікована вірусом, молекули МНС класу І експонуються на мембрані в комплексі з власним (self) антигеном.Мішенню для Т-клітин CD8+ є соматичні та імуно- компетентні клітини, які експонують антигени МНС класу І. Ці молекули відіграють також іншу, не менш важливу роль - розпізнавання нормальних або генетично змінених клітин природними кілерами. Доведено, що саме експресія власних МНС класу І захищає нормальні клітини від дії кілерних клітин, тоді як заміщення власних пептидів на вірусні в комплексі МНС призводить до підвищення лізису клітин-мішеней.
Екзогеннй антиген - це позаклітинний антиген, який був фагоцитований АПК. Процесинг антигену відбувається в ендо- сомах, а утворені фрагменти мають довжину біля 10-20 амінокислотних залишків. Далі процес презентації відбувається так само, як у попередньому випадку, окрім того, що фрагментований пептид зв'язується з молекулами МНС класу ІІ. Комплекс МНС класу ІІ з антигеном розпізнається CD4+-Т-лімфоцитами.
Завдяки можливості вибіркового розпізнавання чужорідних антигенів лише в комплексі зі "своїми", тобто генетично однорідними антигенами МНС (феномен МНС-рестрикції), Т-клітини "наглядають" за змінами в стані клітин організму.
Друга стадія - імунорегуляція - характеризується клональною проліферацією, диференціацією імунокомпетентних клітин, спрямованих на певний антиген, і дією імунорегуляторних медіаторів. На поверхні Т- та В-лімфоцитів є індивідуальні антиге- нозв'язувальні рецептори, завдяки яким вони здатні вибірково розпізнавати та специфічно взаємодіяти з чужорідним антигеном (антигенними детермінантами, або епітопами) за рахунок комплементарності (стеричної спорідненості). Спорідненість просторових конфігурацій обумовлює взаємодію антигенних детермінант вірусу з рецепторами Т- та В-лімфоцитів.
У результаті в лімфоцитах активуються метаболічні процеси і клітини починають проліферувати в клони абсолютно ідентичних клітин, диференціюватись у зрілі клітини, що виконують певні ефекторні функції. Згідно із клонально-селекційною теорією Ф. Бернета (1971), кожному клону імунокомпетентних лімфоцитів притаманний унікальний специфічний рецептор, який здатний взаємодіяти тільки з певною антигенною детермінантою.Регуляція імунної відповіді здійснюється популяцією Т-лімфоцитів, які сприяють установленню напруженого імунітету. Ці клітини розподіляються на дві субпопуляції: CD4 Т-хелпери, CD8 Т-супресори й цитотоксичні. Також виділяють хелпери типів І та ІІ. Т-хелпери типу І беруть участь переважно в розвитку клітинного імунітету, реакціях гіперчут- ливості сповільненого типу й цитотоксичних реакціях; типу ІІ - у розвитку гуморального імунітету й реакціях гіперчутливості негайного типу. Дія Т-хелперів реалізується через продукування регуляторних цитокінів (табл. 4.4).
Таблиця 4.4
Участь Т-хелперів у розвитку противірусного імунітету
| Субпопуляція Т-хелперів | Цитокіни | Ланки імунітету |
| Т-хелпери типу І | ІНФ-у, ІЛ-2, 3; ФНП, ГМ-КСФ | Активація клітинної ланки (знищення внутрішньоклітинного збудника) |
| Т- хелпери типу ІІ | ІЛ-3, 4, 5, 6, 9, 10, 13; ГМ-КСФ | Активація гуморальної ланки (знищення позаклітинного збудника) |
Третя стадія - ефекторна - активація ефекторних клітин, у результаті чого відбувається виділення ефекторних медіаторів, розвиток клітинних реакцій і утворення специфічних циркулюючих антитіл.
Специфічне розпізнавання антигену антитілами й цитотоксичними Т-лімфоцитами зазвичай ініціює потужні неспецифічні ефекторні механізми, які контролюють і знищують інфекційні агенти за допомогою системи комплементу (комплементозалеж- ний лізис), залучення клітин, які відповідають за запальні процеси, фагоцитозу, руйнування клітин (антитілозалежна цито- токсичність клітин).
Якщо вірус перебуває під контролем імунної системи або виводиться з організму, то імунна відповідь поступово згасає. Якщо ж вірус персистує у високих титрах, то результатом є імунопатологія, а персистенція на низькому рівні допомагає підтримувати протективну імунологічну пам'ять.Четверта стадія - стадія імунологічної пам'яті, завдяки якій організм набуває здатності швидко реагувати на повторний контакт зі збудником. Вона характерна для клітинної й гуморальної ланок і залежить від формування дочірніх Т- та В-клітин. Ця форма імунної відповіді пов'язана з накопиченням клону довго- існуючих клітин пам'яті, здатних розпізнавати антиген і прискорено та посилено реагувати на нього. В організмі постійно відбувається ресинтез специфічних імуноглобулінів. Імунологічна пам'ять, особливо Т-клітин, дуже стійка й може зберігатись багато років.
Тривалість постінфекційного імунітету залежить від властивостей збудника, інфікуючої дози, стану імунної системи, генотипу, віку та інших факторів. Імунітет може бути короткочасним (грип), достатньо тривалим (червона висипка) або довічним (кір, поліомієліт).
Основними ланками специфічного набутого імунітету є:
1) опосередкований антитілами гуморальний імунітет,
що залежить від В-лімфоцитів лімфоїдної системи;
2) клітинний імунітет, опосередкований Т-лімфоцитами.
4.5.
Еще по теме Специфічний або набутий противірусний імунітет:
- Андрійчук О. М.. Вірусні інфекції людини та тварин: епідеміологія, патогенез, особливості противірусного імунітету, терапія та профілактика : навч. посіб. / О. М. Андрійчук, Г. В. Коротєєва, О. В. Молчанець, А. В. Харіна. - К. : Видавничо-поліграфічний центр "Київський університет',2014. - 415 с., 2014
- Наступна це система імунітету, захисту.
- Специфічна імунопрофілактика туберкульозу
- Клітинна ланка імунітету
- Гуморальна ланка імунітету
- Пухлина, або бластома
- Непрохідність центральної артерії сітківки або її гілок.
- ГОСТРИЙ ТОНЗИЛІТ АБО АНГІНА
- Сільська місцевість: відсутні або важкодоступні сильнодіючі знеболювальні
- Школа класичного або адміністративно-функціонального управління (А. Файоль, М. Вебер)
- Наука управління або кількісний підхід (1950 — по теперішній час)
- Психологічна характеристика студентського віку як пізньої юності або ранньої дорослості.
- Кардіоміопатії — це первинні захворювання серця невідомої етіології. За класифікацією ВООЗ, їх поділяють на 3 групи: гіпертрофічні, дилатаційні, або застійні, та рестриктивні.
- ГЛОМЕРУЛОНЕФРИТ У ДІТЕЙ. ГОСТРА НИРКОВА НЕДОСТАТНІСТЬ. ХРОНІЧНА НИРКОВА НЕДОСТАТНІСТЬ, АБО ХРОНІЧНА ХВОРОБА НИРОК
- Роль факторів природної резистентності при вірусних інфекціях
- Загальна концепція імунобіології відображає коеволюційну рівновагу між імунною системою та вірусами з метою забезпечення виживання обох: і вірусу, і хазяїна.
- ВІЛ-інфекції
- ЗАПИТАННЯ ДЛЯ САМОКОНТРОЛЮ