Свойства и функции белка предшественника амилоидного пептида (АРР)
АРР представляет собой конститутивный трансмембранный белок, состоящий из 695-770 аминокислот. Он экспрессируется в различных клетках и тканях животных разного уровня эволюционного развития (Johnstone et al., 1991) и принадлежит к большому эволюционно консервативному семейству белков, которое содержит 16 известных на сегодняшний день ортологов (Coulson et al., 2000; Eggert et al., 2004).
В клетках млекопитающих описаны два гомолога АРР, так называемые АРР-подоб- ные белки APLP1 и APLP2 (Jacobsen, Iverfeldt, 2009). В настоящее время известны три изоформы APP (АРР770, АРР751 и АРР695), являющиеся продуктами альтернативного сплайсинга его пре-мРНК и отличающиеся по степени их гликозилирования (рис. 2). APLP1 и APLP2 также подвергаются гликозилированию, но точные сайты этих посттрансляционных модификаций пока не определены (Eggert et al., 2004). Большая часть молекулы АРР (N-концевой фрагмент) находится во внеклеточном пространстве или повернута внутрь везикулярных клеточных органелл, в то время как короткий С-концевой фрагмент молекулы находится в цитоплазме клеток. В нейронах присутствует, в основном, более короткая форма АРР695, которая лишена на своем внеклеточном участке домена ингибитора сериновых протеаз типа Ку- нитца (KPI, kunitz ptrotease inhibitor) и ОХ-2-домена, роль которых до сих пор не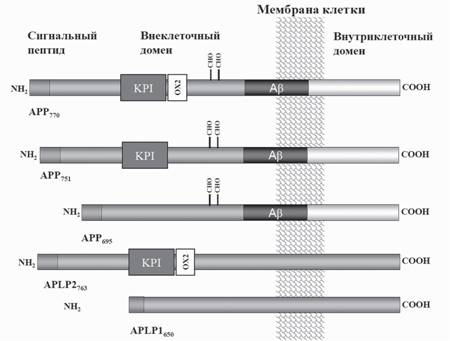
Рис. 2. Схема строения основных изоформ молекулы АРР и расположения участков их протеолитического расщепления
Предшественник амилоидного пептида - АРР, представлен тремя изоформами, содержащими 695, 751 и 770 аминокислотных остатков и отличающимися наличием KPI и/или OX2 доменов. Два АРР-подобных белка, APLP1 и APLP2, содержат 650 и 763 аминокислоты, соответственно, и также различаются наличием KPI и OX2 доменов.
СНО - сайты гликозилирования АРРясна, хотя имеются данные о том, что экспрессия KPI-домена связана с митохондриальной функцией клеток (Chua et al., 2013). В коре мозга человека соотношение различных форм АРР составляет АРР770:АРР751:АРР695=1:10:20 (Tanaka et al., 1989). При развитии БА в ткани мозга наблюдается увеличение содержания АРР770 и АРР751 и снижение АРР695 (Moir et al., 1998). Еще одним различием изоформ АРР является то, что что только нейрональная изоформа АРР695 принимает участие в регуляции экспрессии генов в клетках нерйонального происхождения (Belyaev et al., 2010).
Основным отличием APLP1 и APLP2 от АРР является то, что фрагменты их молекул, образующиеся в результате протеолитического расщепления, и соответствующие последовательности амилоидного пептида в молекуле АРР не образуют амилоидных депозитов. Это объясняется отсутствием в составе этих белков участка (а также соответствующего домена в кодирующих их генах), способствующего формированию амилоидного пептида, характерного для БА (Maloney et al., 2004). В этой же связи интересно отметить, что аминокислотная последовательность АРР грызунов (мышей и крыс) на участке, соответствующем Ар-пептиду, отличается от таковой человека по трем аминокислотам (Arg5 замещен на Gly, Tyr10 на Phe и His13 на Arg) (Fraser et al., 1992), что препятствует образованию типичных Ар-агрегатов в мозге этих животных. Как было показано, наличие гистидина в положении 13 является критическим для способности Ар связывать ионы цинка, необходимые для агрегации молекул пептида и образования фибрилл (Bush, Tanzi, 2002). Именно эта аминокислота отсутствует в данном положении в молекуле Ар крыс и мышей.
Согласно имеющимся литературным данным, АРР обладает нейротрофическими и нейропротекторными свойствами, модулирует рост нервных окончаний и синаптогенез, участвует в обеспечении возбудимости нервных клеток и синаптической пластичности, а также в процессах обучения и памяти (см.
обзоры: Mattson, 1999; Nalivaeva, Turner, 2013). В частности, в процессе развития мозга АРР необходим для правильной миграции нейрональных клеток-предшественников в кортикальную пластину (Young-Pearse et al., 2007). В ходе развития мозга разные изоформы АРР экспрессируются избирательно (Apelt et al., 1997) под действием многочисленных факторов (Lahiri et al., 1995). АРLР белки тоже обладают сина- птогенными свойствами (Thinakaran et al., 1995), а также свойствами регуляторов генной активности и пролиферации клеток. АРР и АРLР2 принимают участие в метаболизме меди (White et al., 1999). В свою очередь, ионы меди модулируют экспрессию гена АРР (Bellingham et al., 2004).Хотя функции АРР еще не полностью установлены, известно, что APP взаимодействует со многими белками и эти взаимодействия играют важную роль в нормальном транспорте, процессинге и внутриклеточных сигнальных каскадах (Aydin et al., 2012). Так, по данным конфокальной микроскопии АРР, колокализу- ется с синаптическими белками, например синаптотагмином (Kohli et al., 2012). Повышение содержания APP в ткани мозга в ходе активного синаптогенеза также свидетельствет о его важной роли в формировании нейрональных связей. Недавно было показано, что АРР необходим для поддержания кальциевого гомеостаза и клеточных осцилляций, которые лежат в основе синаптической передачи (Octave et al., 2013). Растворимая форма АРР, высвобождаемая под действием а-секретазы (sAPPa), также регулирует уровень плотности дендриных шипиков и тем самым принимает участие в процессах обучения и памяти (Jung, Herms, 2012; Tyan et al., 2012). Более того, APP и APLP2 играют важную роль в пространственном обучении и долгосрочном потенциировании (LTP) (Weyer et al., 2012).
Несмотря на то что АРР и его аналоги выполняют столь важные функции в жизнедеятельности клеток и организма, трансгенные мыши, лишенные гена АРР, являются весьма жизнеспособными и фертильными, хотя и обладают меньшим весом, чем мыши дикого типа, и у них наблюдается локомоторный дефицит.
В мозге этих трансгенных мышей к 2 неделям жизни имеет место значительный глиозис, что свидетельствует о нарушении нейрональных функций, вызванных отсутствием в клетках молекул АРР (Zheng et al., 1995). Трансгенные мыши, лишенные гена АРР, а также гена APLP1 или APLP2, тоже являются жизнеспособными, и только тройная мутация APLP1(-/-)/APP(-/-)/APLP2(+/-) приводит к постнатальной гибели животных (Heber et al., 2000).Данные по клонированию и изучению структуры АРР позволяют предположить, что он обладает многими характеристиками, свойственными мембраносвязанным рецепторным молекулам. В частности, работы по реконституции АРР695 и тримерных G-белков свидетельствуют, что АРР может функционировать как рецептор, сопряженный с G-белками (Okamoto et al., 1995). Тем не менее, до сих пор не ясно, что является и существуют ли эндогенные лиганды к рецептору, представленному молекулами АРР, хотя совсем недавно было показано, что представитель одного из классов молекул клеточной адгезии, а именно TAG-1, является реальным кандидатом на роль лиганда АРР (Ma et al., 2008).
Поскольку регуляция экспрессии гена АРР находится под контролем многих факторов, в частности фактора роста TGF-P, цитокина IL-1 и транскирипционного фактора Sp1 (Ge, Lahiri, 2002; Citron et al., 2008), это рассматривается как один из возможных путей направленного контроля экспрессии АРР с целью снижения образования токсичного Ар. Однако, принимая во внимание участие АРР в синап- тогенезе и функционировании нервной системы, данный подход к лечению БА мы представляем неперспективным, поскольку снижение экспрессии АРР и родственных ему АРРL1/2 может повлечь нежелательные побочные эффекты.
Недавно было показано, что микро-РНК (miR), представляющие собой короткие некодирующие РНК, взаимодействющие с молекулами РНК-мишеней и ингибирующими их трансляцию или транспорт, также принимают участие в регуляции экспрессии гена АРР и играют важную роль в развитии нейродегенерации (Salta, De Strooper, 2012). В частности, miR-101 и miR-153 снижают экспрессию APP в культуре нейрональных клеток человека, предполагая их возможное участие в патогенезе БА (Long, Lahiri, 2011; Long et al., 2012). Поскольку микро-РНК играют важную роль в эпигенетической регуляции экспрессии генов, весьма вероятно, что их нормальное функционирование изменяется в ходе патогенеза БА. Данные последних лет свидетельствуют, что существует довольно много форм микро-РНК, которые нарушены при данной патологии и их дальнейшее исследование может иметь большое теоретическое и практическое значение (Van den Hove et al., 2013).
2.2.
Еще по теме Свойства и функции белка предшественника амилоидного пептида (АРР):
- Роль предшественника амилоидного пептида и его метаболитов в патогенезе болезни Альцгеймера
- Протеолитический процессинг предшественника амилоидного пептида
- Свойства амилоидного пептида
- Харктеристика амилоидного пептида
- Основные свойства и характеристики морфогенетического белка кости
- Белки предшественники амилоидов
- Амилоидное (сальное, восковое) перерождение
- Метаболизм Ар-пептидов в нормально функционирующих нейронах
- Результаты собственных исследований. Эксперимент со стромальными клетками-предшественниками костного мозга.
- Нитрозамины и их предшественники.
- 13.2 Свойства конкретного человека и свойства системы
- Очистка рекомбинантного белка (задача 5)
- АТРИАЛЬНЫЙ НАТРИЙУРЕТИЧЕСКИЙ ПЕПТИД (Atrial Natriuretic Peptide (ANP[1-28])
- Получение эксплантационного материала в эксперименте на культуре клеток-предшественников стромальных фибробластов костного мозга.
- Антимикробные пептиды
- Определение R-белка
- Изучение С-реактивного белка при воспалительных заболеваниях
- Нарушение реабсорбции белка.
- Выделение рекомбинантнного белка из биомассы
- Особенности синтеза белка в нейронах