Протеолитический процессинг предшественника амилоидного пептида
Уровень содержания АРР в клетках и тканях регулируется при помощи его направленного протеолиза под действием специфических ферментов, расщепляющих этот трансмембранный белок с образованием ряда растворимых биологически активных продуктов.
В настоящее время выделяют три основные сайта расщепления АРР под действием различных ферментов или белковых комплексов, называемых собирательно a-, P- и у-секретазами. Они способны расщеплять АРР с образованием фрагментов разной длины, обладающих определенными функциональными свойствами (рис. 3). В результате действия а- или Р-секретаз образуются крупные растворимые белки sAPPa и sAPPp, играющие важную роль в процессах пролиферации нервных клеток и развитии нервной ткани (Caille et al., 2004; Turner et al., 2007; Chasseigneaux, Allinquant, 2012), и мембраносвязанные фрагменты С83 и С99. В результате действия у-секретазы на С99 образуется Ар и короткий цитоплазматический фрагмент AICD, играющий важную регуляторную роль (Muller et al., 2008; Beckett et al., 2012), а при действии на С83 - короткий пептид р3 с неизвестными функциями и AICD, который быстро подвергается протеолитическому расщеплению (Beckett et al, 2012; Venugopal et al., 2007). Таким образом, катаболизм АРР может осуществляться как по амилоидогенному пути в результате действия P- и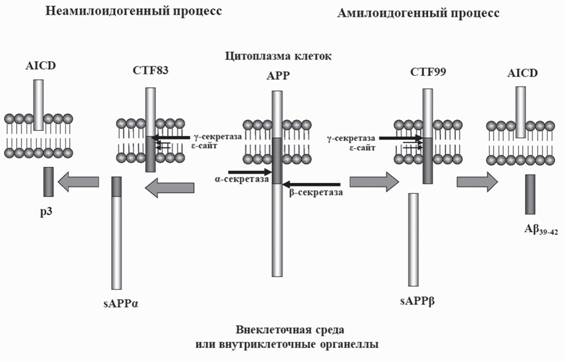
Рис. 3. Амилоидогенный и неамилоидогенный пути расщепления АРР
При неамилоидогенном расщеплении, по которому метаболизирует 95% АРР, молекула этого белка подвергается действию а-секретазы с образованием растворимого sAPPa и мембраносвязанного С-концевого фрагмента С83, из которого под действием у-секретазы образуется AICD и короткий нетоксичный пептид p3. При амилоидогенном пути АРР сначала расщепляется Р-секретазой с образованием растворимого sAPPp и мембраносвязанного С-концевого фрагмента С99, из которого в результате действия у-секретазы образуется Ар и транскрипционно активный AICD
у-секретаз с образованием Ар, так и по неамилоидогенному пути, в котором принимает участие а-секретаза (Hooper, Turner, 2002).
Этот фермент расщепляет АРР внутри последовательности Ар между лизином и лейцином в положении 16 и 17, соответственно, тем самым предотвращая образование Ар пептида. Исходя их этого, регуляция (повышение) активности а-секретазы является одним из возможных подходов для профилактики БА (Lichtenthaler, Haass, 2004; Fahrenholz, 2007). Помимо а-, P- и у-участков расщепления АРР выделяют также сайты, обозначаемые как е- и 5. В то время как s-участок находится в непосредственной близости от места действия у-секретазы, приводя к образованию АРРе, который является субстратом основных секретаз (Lefranc-Jullien et al., 2006), 5-сайт локализован в районе действия Р-секретазы (12 аминокислотных остатков от N-конца АР) и также приводит к образованию пептида, склонного к аггрегации (Eggert et al., 2004). Однако до настоящего времени расщепление АРР по этому пути было обнаружено только в гиппокампальных нейронах (Simons et al., 1996) и не подтверждено другими исследователями. С другой стороны, 5-сайт расположен совсем рядом с недавно обнаруженным участком действия Р-секретазы-1 (ВАСЕ1), которая расщепляет АРР возле Glu11 (относительно N-конца на участке АР) и продукт расщепления образующегося при этом С-концевого фрагмента АРР (АРР89) является субстратом у-секретазы, высвобождающей неамилоидогенный укороченный Ар-пептид (Deng et al., 2013).В настоящее время известно несколько белков, которым приписывают а-секретазную активность. В основном это цинк-зависимые металлопротеиназы, или адамализины, принадлежащие к семейству ADAM (a disentegrin and metalloprotease), в частности ADAM10 и ADAM17 (Hooper, Turner, 2002; Deuss et al., 2008, Vincent, Checler, 2012). ADAM17 также известен как ТАСЕ (TNFa-кон- вертирующий фермент), принимающий участие в секреции фактора некроза опухолей (Allinson et al., 2004). ADAM9, или MDC9, также обладает а-секретазной активностью (Koike et al., 1999). Конститутивный процесс секреции растворимого АРР (sAPPa) под действием а-секретазы (см. рис. 3) происходит на поверхности клеток (Parvathy et al., 1999) и по этому пути происходит расщепление до 90% всего клеточного АРР.
В то же время, регулируемая различными факторами, например, холинергическими агонистами мускарином и карбахолом (Canet-Aviles et al., 2002) или форболовыми эфирами (Zhu et al., 2001) активность а-секретазы локализуется, в основном, в аппарате Гольджи. Активность а-секретазы ингибируется пептид- гидроксаматами, в частности батимастатом (Parvathy et al., 1999). При умеренной оверэкспрессии белка ADAM10 в нейрональных клетках образование Ар снижается и формирования сенильных бляшек не наблюдается. Напротив, экспрессия мутантных молекул ADAM10, не способных к протеолизу АРР, приводит к увеличению числа амилоидных бляшек в мозге трансгенных мышей, экспрессирующих мутантный человеческий АРР (Postina et al., 2004). Трансгенные животные, лишенные гена ADAM10 или ADAM17, являются нежизнеспособными, свидетельствуя о важной роли а-секретазы в жизнедеятельности организма (Sahin et al., 2004).Образуемый в результате действия а-секретазы внеклеточный фрагмент sAPPa обладает не только нейропротекторными свойствами, но также свойствами эпидермального фактора роста (Wehner et al., 2004), а также способствует обучению и памяти (Turner et al., 2007; Chasseigneaux, Allinquant, 2012). Образование sAPPa посредством активации а-секретазы находится под контролем рецепторов, сопряженных с G-белками (Racchi, Govoni, 2003), а также таких факторов, как Epac, регулирующий цАМФ-зависимый обмен гуаниновых нуклеотидов, и ГТФ-аза Rac (Robert et al., 2005). Важную роль в регуляции активности а-секретазы также играют ионы железа (Bodovitz et al., 1995).
Еще один продукт, образующийся при неамилоидогенном расщепления АРР а-секретазой, представляет собой С-концевой фрагмент молекулы АРР (С83), состоящий из 83 аминокислотных остатков и являющийся субстратом у-секретазы (см. рис. 3). При его расщеплении образуется внутриклеточный С-концевой фрагмент АРР (AICD) и короткий пептид массой около 3 кДа (р3). Свойства р3 пептидов, состоящих из аминокислотных остатков 17-40 или 17-42 последовательности Ар, пока не известны.
Согласно имеющимся данным, AICD, образующийся по а-секретазному пути, быстро подвергается протеолитическому расщеплению и не вносит существенный вклад в регуляцию генов (для обзора см. Beckett et al., 2012). В целом, эта часть процесса протеолиза АРР, запускаемого а-секретазой, считается неамилоидогенной и не играющей существенной роли в патологии БА (Kimberly et al., 2000).Образование Ар требует участия двух ферментов, именуемых P- и у-секретазами (для обзора см. Mattson, 2004). В ходе многолетних исследований было показано, что фермент, обладающий Р-секретазной активностью, является новым представителем семейства трансмембранных аспартатных протеаз. Этот фермент, получивший название P-site APP-cleaving enzyme, или BACE1 (другие названия Asp2 и ме- мапсин), инициирует образование Ар (Vassar, 2004). В литературе описан еще один фермент, гомологичный ВАСЕ1 и получивший название ВАСЕ2. Этот фермент также обладает Р-секретазной активностью по отношению к АРР (Bennett et al., 2000).
BACE1 является трансмембранным белком, со стоящим из 501 аминокислотного остатка. Активная форма этого фермента, подобно другим аспартатным протеазам, таким как пепсин и катепсин Д, образуется путем отщепления 23 аминокислотных остатков от неактивной молекулы белка-предшественника. Однако ВАСЕ1 отличается от указанных протеаз по вторичной структуре (Hong et al., 2000). В литературе описано несколько вариантов посттрансляционных модификаций этого фермента, которые вносят разный вклад в патогенез БА (для обзора см. Holsinger et al., 2013). BACE1 расщепляет АРР между метионином и аспартатом в положении 671 и 672, соответственно, в молекуле APP, состоящей из 770 аминокислот (Haass et al., 1992). ВАСЕ2 также способен расщеплять АРР на этом участке, а также ближе к С-конце- вому участку молекулы - в районе тирозина-10 и глутамата-11 внутри последовательности Ар, что ведет к образованию более короткого варианта Ар (Farzan et al., 2000). Как было показано совсем недавно, ВАСЕ1 также расщепляет АРР на этом участке и, по мнению авторов, в нормально функционирующем мозге этот процесс преобладает.
Однако наличие мутаций в молекуле АРР или другие патологические воздействия приводят к предпочтительному расщеплению АРР с образованием токсичного Ар (Deng et al., 2013). Как было показано в наших исследованиях, нейрональная изоформа АРР695 расщепляется преимущественно по Р-секретазному пути с образованием Ар и транскрипционно активного AICD (Belyaev et al., 2010).С терапевтической точки зрения, ВАСЕ1 представляет собой довольно перспективный фермент, поскольку мыши, лишенные гена этого белка, не образуют Ар и являются жизнеспособными (Luo et al., 2001). В связи с этим было предложено несколько подходов для создания эффективных ингибиторов Р-секретазной активности у человека в основном с использованием пептидомиметиков, являющихся не превращаемыми аналогами субстратов ВАСЕ. Другой подход предусматривает создание молекул, распознающих BACE1 мРНК и снижающих его экспрессию. Последние включают в себя антисмысловые олигонуклеотиды, каталитические нуклеиновые кислоты - рибозимы и дезоксирибозимы, а также небольшие интерфе- ририрующие молекулы РНК (siRNAs) (Nawrot, 2004). Тем не менее, недавно были обнаружены новые эндогенные субстраты ВАСЕ1, что заставляет относиться к созданию ингибиторов этого фермента с определенной осторожностью. В частности, было показано, что ВАСЕ1 играет важную роль в функицонироавнии сетчатки и подавление ее активности приводит к ретинопатии (Cai et al., 2012).
Весьма неожиданным, но с биохимической точки зрения предсказуемым, было обнаружение того, что активность Р-секретазы регулируется конститутивными клеточными белками прионами PrPc, модифицированные формы которых (PrPsc) являются патогенным фактором развития болезней Крейцфельда-Якоба и коровьего бешенства (Parkin et al., 2007). Это указывает на наличие связи между метаболизмом Ар и PrPc. В нормальном мозге последние регулируют активность Р-секретазы, препятствуя накоплению Ар. Нарушение синтеза PrPc или его конформации в случае болезней Крейцфельда-Якоба или коровьего бешенства ведет к накоплению амилоидного пептида, что также является характерным признаком данных заболеваний (Hooper, Turner, 2008).
С другой стороны, мутации АРР, приводящие к развитию ювенильной формы БА, также нарушают процесс регуляции ВАСЕ1 при участии PrPc, что приводит к усугублению патологии (Griffiths et al., 2011).Как уже упоминалось выше, вторым ферментом, принимающим участие в образовании Ар, является у-секретаза, представляющая собой сложный комплекс из нескольких белковых молекул. До сих пор нет окончательного мнения относительно состава и строения у-секретазы, однако общепринято, что это многокомпонентный комплекс, состоящий из белков пресенилинов 1 и 2 (PS1 и PS2), никастрина (NCT) и белков Aph-1 и Pen-2 (Shirotani et al., 2004; для обзора см. Wolfe, 2012). Помимо протеолитического расщепления С-концевого фрагмента АРР у-секретазный комплекс принимает участие в недавно открытом, но еще мало понятном пути регулируемого трансмембранного протеолиза (protein RIPing) (Heldin, Ericsson, 2001). Компонент у-секретазы PS1 также принимает участие в протеолитическом расщеплении сигнальных молекул Notch, играющих важную роль в развитии ЦНС (для обзора см. Lathia et al., 2008).
Белки пресенилины были открыты в ходе скрининга генетических мутаций, ведущих к формированию ювенальной формы БА (Sherrington et al., 1995; Rogaev et al., 1995). Пресенилины вместе с протеазами сигнальных пептидов (SPP) представляют собой новый класс протеаз, являющихся интегральными мембранными белками, в основе деятельности которых лежат каталитические свойства классических аспартатных протеиназ, таких как пепсин, ренин и катепсин Д. Характерной чертой пресенилинов и протеаз сигнальных пептидов является их способность расщеплять полипептидные субстраты внутри трансмембранного домена. Это требует присутствия в каталитическом участке молекул этих ферментов двух соседствующих остатков аспартата и консервативного Про-Ала-Лей мотива в С-концевой части их молекул. Именно пресенилины, как было подтверждено рядом авторов, являются каталитическими субъединицами мультипротеинового комплекса, который проявляет у-секретазную активность (De Strooper, 2003). Клеточные и молекулярно-биологические исследования показали, что пресенилины и их гомологи имеют 9 трансмембранных доменов, а недавно полученная кристаллическая структура этого белка представляет платформу для выяснения субстратной специфичности этих ферментов и создания их ингибиторов (Wolfe, 2013).
В физиологических условиях APP может расщепляться не только внутри его трансмембранного домена, но также на цитоплазматическом участке, близком к плазматической мембране, с образованием С-концевого фрагмента С50 и более длинного фрагмента АРРе, который, в свою очередь, является субстратом а- и Р-секретаз, а также у-секретазы (Lefranc-Jullien et al., 2006). Как было показано, расщепление АРР на у- и е-участках представляет собой два независимых процесса, которые регулируются разными механизмами. При БА имеет место достоверное снижение е-секретазной активности (Kametani, 2008).
В результате действия у-секретазы на С99 фрагмент АРР образуется набор Ap пептидов, содержащих 40, 42 или 43 аминокислотных остатка, что предполагает тем самым существование нескольких сайтов расщепления. Тем не менее, AP40 образуется в наибольшей концентрации, в то время как AP42 является более токсичным, менее растворимым и с большей готовностью образует конгломераты (El-Agnaf et al., 2000). Исследования мембранной топологии убедительно показали, что мутации АРР в районе трансмембранного домена драматически изменяют соотношение AP40/AP42 в сторону образования высоких концентраций более токсичной формы AP42 (Lichtenthaler et al., 1999). Механизм, в результате которого мутации в молекуле АРР и пресенилинов ведут к гиперпродукции более длинных изоформ Ap, пока еще не известен, но в данном направлении ведутся довольно многочисленные исследования (Van Dijk et al., 2004).
Недано было показано, что активность у-секретазы находится под контролем Р-аррестина 2, принадлежащего к классу соединений, контролирующих многочисленные функции рецепторов, связанных с G-белками. Так, при развитии БА аррестин 2 непосредственно связывается с Aph-1, приводя к предпочтительному распределению у-секретазного комплекса в липидные домены клеточных мембран, обогащенные сфинголипидами и холестерином, где происходит амилоидогенное расщепление АРР и образование Ар. Это ведет к повышению концентрации последнего и усугублению патологии (Thathiah et al., 2013).
Несмотря на то что создание ингибиторов у-секретазного комплекса для предотвращения формирования Ap и развития БА является весьма привлекательным теоретическим подходом, он имеет определенные ограничения, поскольку мыши, лишенные обеих аллелей гена PS-1, погибают, демонстрируя существенную патологию развития (Shen et al., 1997). Более того, ингибиторы у-секретаз также приводят к патологии развития вследствие нарушения сигнальных процессов, осуществляемых посредством рецепторов Notch (Kopan, Ilagan, 2009). Это диктует необходимость направленного поиска способов ограничения протеолиза АРР при участии у-секретазы, которые бы не затрагивали процессинг Notch рецепторов (Wolfe, 2012).
В апоптотических клетках и в мозге больных деменцией альцгеймеровского типа АРР помимо секретаз может также подвергаться альтернативному протеолизу под действием каспазы-3 и каспазы-8 (Gunyuzlu et al., 2000). Расщепление под действием каспаз происходит внутри цитоплазматического домена молекулы АРР в районе аспартата 664 (в молекуле АРР, состоящей из 695 аминокислотных остатков). Продукты протеолиза АРР каспазами являются очень токсичными. В нормально функционирующих клетках есть механизм защиты АРР от действия каспаз посредством фосфорилирования остатка аминокислоты треонина в положении 668 молекулы АРР. Изменение характера фосфорилирования АРР и повышение его чувствительности к каспазам рассматривается как один из факторов патогенеза БА (Taru et el., 2004). При определенных условиях каспазы также расшепляют цитоплазматический фрагмент АРР (AICD), который принимает участие в регуляции экспрессии ряда генов, о чем более подробно будет изложено в последующих разделах данной главы.
Протеолитический процессинг АРР, как было показано в многочисленных исследованиях, происходит в так называемых липидных рафтах, которые представляют собой специализированные домены клеточных мембран, обогащенные холестерином и ганглиозидами, в частности GM1 (для обзора см. Hicks et al., 2012). Эти домены играют важную роль в функционировании клеток, обеспечивая такие функции, как связывание лигандов (включая вирусы и олигомеры АР) и передача сигналов, развитие аксонов и поддержание целостности синаптических окончаний. Именно на этих участках происходит взаимодействие АРР и ВАСЕ1 и инициируется процесс образования Ap. Нарушение целостности этих мембранных структур приводит к клеточной гибели и нейродегенерации. В связи с этим модификации липидного состава нервной ткани являются одними из важных факторов, влияющими на амилоидный метаболизм и развитие БА (Walter, van Echten-Deckert, 2013).
Помимо процессов расщепления АРР, которые вносят свой вклад в патогенез БА, существенную роль в определении амилоидного метаболизма также играют процессы транскрипции и трансляции гена АРР, которые приводят к формировании его изоформ, а также генов его основных секретаз. В недавних исследованиях было показано наличие корреляции между уровнем накопления в нерастворимых белковых комплексах, характерных для БА и других нейродегенеративных заболеваний, коротких ядерных рибонуклеобелков (U1 snRNP), принимающих участие в сплайсинге молекул мРНК, и уровнем развития когнитивного дефицита на стадии MCI и БА. Авторы высказывают прпедположение, что дефицит активности этих U1 snRNP комплексов приводит к нарушению нормального сплайсинга мРНК и накоплению неспецифических для нервных клеток изоформ белков, в частности АРР и ВАСЕ1 (Bai et al., 2013). С другой стороны, становится все более очевидным, что короткие некодириующие микро-РНК (miR) также обеспечивают контроль за синтезом АРР и других белков, вовлеченных в патогеенез БА, и эти процессы могут быть нарушены или изменены при патогенезе БА (Lau et al., 2013). Так, в частности, была показана роль miR-29a/b в синтезе ВАСЕ1 и расщеплении АРР (Herbert et al., 2008). В целом, молекулы miR в настоящее время рассматриваются как возможные маркеры БА (Kiko et al., 2014).
2.3.
Еще по теме Протеолитический процессинг предшественника амилоидного пептида:
- 1.7.3. Болезнь Альцгеймера
- IY-2.1. СТРУКТУРА НЕЙРОДЕГЕНЕРАТИВНЫХ ПАТОЛОГИЙ
- Выявление пусковых факторов амилоидоза нейронов в начальной стадии болезни Альцгеймера
- Образование конформационно модифицированных наноамилоидов с разной архитектурой
- Роль предшественника амилоидного пептида и его метаболитов в патогенезе болезни Альцгеймера
- Свойства и функции белка предшественника амилоидного пептида (АРР)
- Протеолитический процессинг предшественника амилоидного пептида
- Участие сфинголипидов в амилоидогенезе