Белки предшественники амилоидов
АРР представляет собой крупный конститутивный трансмембранный белок. Существуют три изоформы АРР, отличающиеся по молекулярному весу и представленные в разных соотношениях в нейроне (Tanaka et al., 1989).
АРР входит в семейство интегральных мембранных белков, в котором представлены также APLP1 и APLP2, которые обладают высокой степенью гомологии с АРР, но не содержат последовательности Ар (De Strooper, Annaert, 2000; Wasco et al., 1992, 1993). Хотя клеточные функции семейства АРР до сих пор остаются не вполне понятными, существуют данные о том, что все три белка вовлечены в аксонный транспорт и необходимы для нормального развития нервной системы млекопитающих (Hardy, Selkoe, 2002). Он необходим для клеточной адгезии, пролиферации клеток и межклеточных взаимодействий. При этом в мембраносвязанном состоянии АРР могут выполнять сигнальную рецепторную функцию в процессе нейрогенеза (De Strooper, Annaert, 2001; Wilquet, De Strooper, 2004) и принимать участие в образовании и поддержании синапсов и увеличении синаптической плотности (Moya et al., 1994; Roch et al., 1994). АРР в клетках локализован как в наружных плазматических, так и внутриклеточных мембранах, и его амилоидогенный протеолиз происходит преимущественно в компартментах мембран, богатых холестерином и ганглиозитом GM1 (так называемых липидных рафтах) (Cordy et al., 2003; Mattson, 1997).Белки семейства АРР (695-770 АК) состоят из гидрофильного N-концевого внеклеточного домена, гидрофобного трансмембранного домена и С-концевого цитоплазматического домена (Kang et al., 1987) (рис. 1). Сравнение ДНК человека и других млекопитающих показывает высокую степень консервативности гена АРР: установлена 100-процентная идентичность АРР-695 из мозга человека и обезьян. В нейронах ЦНС доминирует изоформа АРР-695, а формы АРР-770 и АРР-751 присутствуют в следовых количествах (Stanton, Coetzee, 2004; Ещенко, 2004).
АРР синтезируется и гликозилируется в эндоплазматическом ретикулуме, затем переносится в комплекс Гольджи для созревания перед транспортом к клеточной поверхности (Molinari et al., 2004). Был идентифицирован ряд молекулярных шаперонов, которые принимают участие в данном процессе, в том числе при высокой температуре (HtrA) (Molinari et al., 2004). Представленные данные косвенно свидетельствуют об участии АРР в энергетических процессах нейронов. В норме биологическая функция АРР состоит в перемещении клеточного материала внутри нейронов, регуляции синапсогенеза и нейрональной трансмиссии, а также в обеспечении внеклеточной адгезии и миграции (Priller et al., 2006). Этот трансмембранный белок играет важную роль в росте нейрона, его выживании и в восстановлении после повреждений (Turner et al., 2003). Если молекула АРР сразу после синтеза активно фосфорилируется (Tarr et al., 2002), то дальнейший процессинг происходит при участии а-секретазы по неамилоидному пути в направлении от центра к периферии (см. рис.1). Фосфорилированный АРР продвигается к мембране, где от него отщепляются фосфаты (Alonso et al., 2006; von Bergen et al., 2000). При нормальных условиях около 90% АРР расщепляется по неамилоидному пути (Steiner et al., 2008; На- ливаева и др., 2008). Активация гена DYRK1A способствует фосфорилированию АРР по Thr-668 in vitro и in vivo, которое может приводить к образованию фрагментов. При недостаточном фосфорилировании процессинг молекул АРР переключается при участии Р-секретазы на амилоидный путь. Финальное расщепление АРР у-секретазой при амилоидном процессинге может происходить по нескольким участкам. В результате образуются пептиды длиной 39-43 аминокислотных остатков, но преимущественными вариантами являются Ар40 и Ар42 (Steiner et al., 2008). Продукты расщепления АРР а-секретазой (не амилоидный путь) также подвергаются дальнейшему протеолизу под действием у-секретазы. Однако этот процесс не ведет к образованию амилоидного пептида. Важно отметить, что образующиеся в результате процессинга а- и у-секретаз крупные и растворимые фрагменты АРР выполняют защитные функции: имеют нейротрофические свойства, являются ней-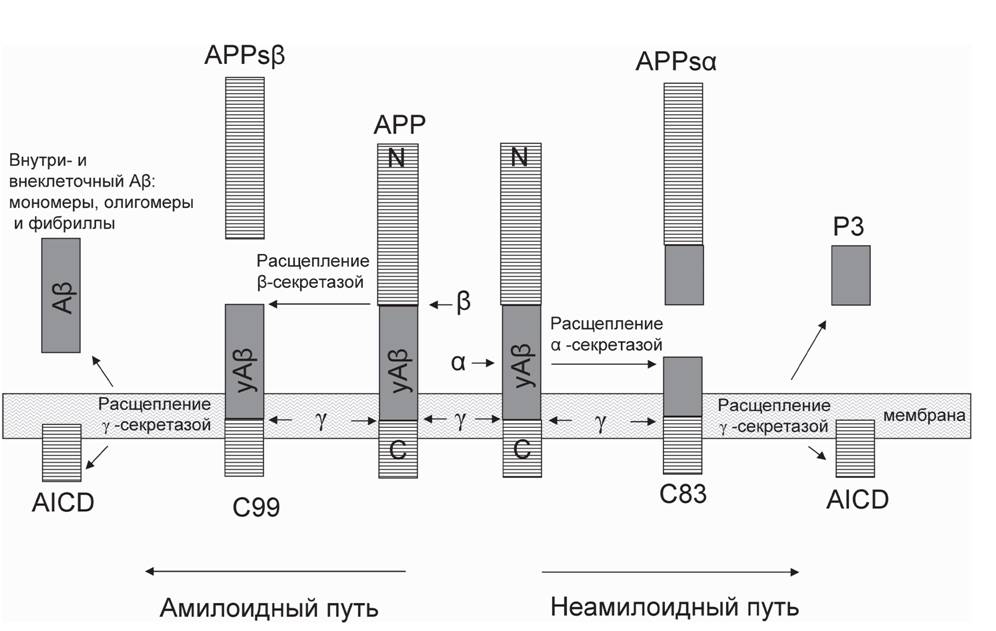
Рис. 1. Амилоидный и неамилоидный пути расщепления белка-предшественника амилоидного пептида. уАр - участки будущих молекул Ap в структуре белковой молекулы APP
ропротекторными и необходимы для нормального функционирования нейронов, в частности для синаптогенеза и для формирования памяти (Turner et al., 2003, 2007). Как при амилоидном, так и неамилоидном процессинге АРР действие у-секретазы ведет к образованию внутриклеточного концевого фрагмента (AICD), который является фактором транскрипции и, вероятно, регулирует уровень экспрессии как самого АРР по принципу обратной связи, так и амилоид-деградирующего фермента неприлизина (Belyaev et al., 2009; Pardossi-Piquard et al., 2005).
6.
Еще по теме Белки предшественники амилоидов:
- Характеристика сывороточного амилоида А
- Роль предшественника амилоидного пептида и его метаболитов в патогенезе болезни Альцгеймера
- Роль в-амилоида в патогенезе БА
- Локализация амилоида. Вопросы патогенеза.
- в-амилоид и периферические нарушения: вклад в нейродегенеративный процесс
- Результаты собственных исследований. Эксперимент со стромальными клетками-предшественниками костного мозга.
- Амилоид-деградирующие ферменты
- Нитрозамины и их предшественники.
- Протеолитический процессинг предшественника амилоидного пептида
- Роль амилоида-р в болезни Альцгеймера
- Регуляция свойств амилоид-деградирующих ферментов
- Свойства и функции белка предшественника амилоидного пептида (АРР)
- Измерение концентрации амилоида А и CA125 методом иммуноферментного анализа
- Белки
- Белки острой фазы
- Глава 5 Мембранные белки
- Белки теплового шока и БА
- Получение эксплантационного материала в эксперименте на культуре клеток-предшественников стромальных фибробластов костного мозга.