Механизмы ингибирования синтеза дигидрофолиевой кислоты сульфаниламидными препаратами
Открытие фолиевой кислоты позволило глубже понять механизм действия сульфаниламидных препаратов в качестве антагонистов ПАБ. Птероилглутаминовая кислота — ярко-желтое вещество, выделенное из печени, дрожжей, листьев и бактерий, предупреждающее малокровие у позвоночных, было названо фолиевой кислотой [Waller et al., 1948].
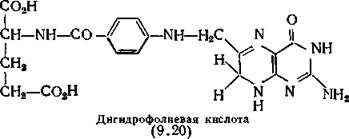
Структура этого соединения, подтвержденная синтезом, представлена формулой (9.20), но без двух атомов водорода в положениях 7 и 8, отщепляющихся от молекулы дигидрофолиевой кислоты при выделении. Молекула дигидрофилиевой кислоты (9.20) состоит из трех фрагментов: остатков глутаминовой кислоты, ПАБ и 2-амино- €-метил-4-оксоптеридина.Бактерии используют ПАБ только для превращения ее в 7,8-дигидрофолиевую кислоту [Griffin, Brown, 1964]. Так бактерии Е. coli участвуют в конденсации ПАБ или пара- амино- бензоилглутаминовой кислоты с 2-амино-4-гидроксиметил-7,8- оксоптеридином (9.21) (точнее его 6-пирофосфатом), что приводит к образованию дигидроптероиновой (или дигидрофолиевой) кислоты [Jaenicke, Chan, 1960]. Эту реакцию катализирует фермент дигидрофолатсинтетаза, которую конкурентно ингибируют сульфаниламидные препараты. Из бактерий Lactobacillus planta- rum [Shiota et al., 1969a] были выделены кв а чистых фермента, ответственные за этот синтез. Первый из них катализирует этерификацию птеридина (9.21) в его 6-пирофосфорил производное. Второй оказался дигидрофолатсинтетазой, обнаруженной ранее G. Brown (1962). Второй фермент также был выделен из нескольких штаммов Pneumococcus, причем было установлено, что его ОММ равна 90 000, а коферментами служат АТФ и Mg2+ [Ortiz, 1970].
ЗО
Таблица 9.2. Конкурентное и неконкурентное ингибирование ПАБ [Lam- pen, Jones, 1946]
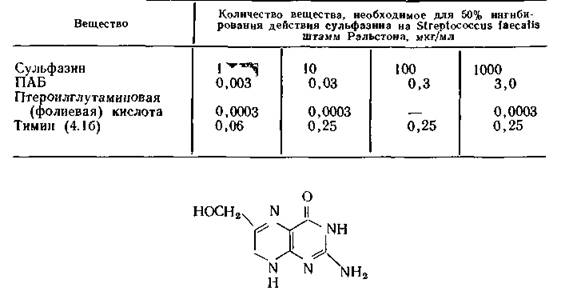
Производное птеридина — промежуточное соединение в биосинтезе фолиевой кислоты
(9.21)
Сульфаниламиды не ингибируют действие птероиновой и фолиевой кислот (а также их 7,8-дегидропроизводных).
Наиболее четко это было показано на примере нескольких штаммои бактерий, способных поглощать фолиевую кислоту (например,, штаммы Streptococcus faecalis и некоторые представители Lactobacilli). Эти бактерии не патогенны для человека. В табл. 9.2' показана прямая корреляция между количеством сульфазина,. необходимым для угнетения роста бактерий Streptococcus faecalis (штамм Ральстона), и количеством ПАБ, требуемым для снятия ингибирующего действия. В отличие от этого количество- фолиевой кислоты, необходимое для снятия ингибирующего- эффекта, остается постоянным, независимо от количества использованного сульфазина. Это свидетельствует о том, что суль- фазин нарушает синтез фолиевой кислоты из ПАБ, но не влияет на ее дальнейшие превращения в отличие, например, от метотрексата. Аналогично сульфаниламиды угнетают рост L. ага- binosus (которые нуждаются во внешнем источнике ПАБ),, и это угнетение конкурентно снимается ПАБ и неконкурентно — посредством фолиевой кислоты. Количество фолиевой кислоты (определяемое по разрушению бактерий и титрованием гомогенизированной бактериальной культуры бактериями L. casei, чувствительными к фолиевой кислоте, но не к ПАБ), которое вырабатывается этими микроорганизмами, обычно пропорционально количеству ПАБ в среде. Однако, если в среде присутствует и сульфаниламид, количество фолиевой кислоты уменьшается пропорционально его концентрации (в 10000-кратном интервале ее измерения) [Nimmo-Smith, Lascalles, Woods, 1948].Образование фолиевой кислоты в культуре Е. coli также ингибируется сульфаниламидами [Muller, 1944].
Подобное ингибирование ферментов сульфаниламидами, по- видимому, составляет основу терапевтического действия этих препаратов. Однако в некоторых экспериментах взаимодействие стрептоцида с птеридиновым интермедиатом (9.21) приводило к образованию устойчивого комплекса. Так, например, при выдерживании фермента Брауна в термостате в течение 2 ч в бесклеточной системе, содержащей стрептоцид, ингибирующее действие последнего не удалось снять с помощью ПАБ; в растущих клетках такой эффект не наблюдали [G.
М. Brown, 1962]. Для изучения этого явления использовали фолатсинтезирующие ферменты из Е. coli: под их действием сульфаметоксазол (9.11) был превращен в аналог птероиновой кислоты — N'-З- (5-метилизок- сазолил) -N4- (7,8-дигидро-6-птероилметил) сульфаниламид, выделенный с помощью хроматографии. Полученное вещество оказалось идентичным синтетическому образцу [Bock et al., 1974].Из всех реакций, катализируемых фолиевыми кислотами, синтез тимина наиболее чувствителен к недостатку этих кислот (разд. 9.3.2), хотя в некоторых микроорганизмах первым ингибируется синтез пуринов. Из данных, приведенных в табл. 9.2, можно видеть, что тимин способен подавлять противострепто- кокковую активность сульфазина, причем его действие является почти неконкурентным. Не удивительно, что при этом требуется большое количество тимина, так как тимин — не катализатор (подобно ПАБ и фолиевой кислоте), а компонент клетки и потребность в нем увеличивается по мере роста клетки.
Действие лекарственных препаратов — антагонистов ПАБ на большинство видов патогенных бактерий и простейших не снимается с помощью фолиевой кислоты и ее производных, так как они не способны проникать в эти микроорганизмы.
9.3.2.
Еще по теме Механизмы ингибирования синтеза дигидрофолиевой кислоты сульфаниламидными препаратами:
- Характеристика препарата урсодезоксихолевой кислоты (УДХК)
- Антифибринолитический препарат (Транексамовая кислота) в лечении МКПП
- Б. Механизм связывания акридинов и их аналогов с нуклеиновыми кислотами.
- А. Механизм действия препаратов мышьяка при лечении сифилиса.
- Общие сведения о патогенетических механизмах женского бесплодияпри разновременном воздействии компонентов синтеза карбамидно-формальдегидной смолы
- Б. Ингибирование ДНК- и РНК-полимеразы.
- Общая характеристика социальной защиты и её правовых механизмов для лиц в клинических исследованиях лекарственных препаратов для медицинского применения
- Методические рекомендации по постановке тестов ингибирования роста бактерий, выделенных в ветеринарных лабораториях при диагностике болезней животных
- Роль фолиевой кислоты и других птеридинов
- 9.3. Антагонисты фолиевой кислоты
- Летальный синтез
- Глава III. Некоторые аспекты биосинтеза жирных кислот у грибов
- Жирные кислоты
- Отравление кислотами.
- Другие примеры летального синтеза.
- Жирные кислоты грибных фосфолипидов