Б. Ингибирование ДНК- и РНК-полимеразы.
Два факта ука-
Таблица 10.9. Выделение ионов водорода из бактерий Bacillus beltus, вызванное адсорбцией катионов кристаллического фиолетового1 (pH измеря- ли стеклянным электродом) [McCalla, 1941]
| pH бактериальной суспензии (1) | pH раствора кристаллического фиолетового (2) | pH смеси (1) и (2) | Снижение pH, вызываемое адсорбцией |
| 8,5 | 8,5 | 6,6 | 1,9 |
| 6,5 | 6,3 | 4,8 | 1.7 |
| 5,5 | 5,5 | 3,8 | 1,7 |
| 5,0 | 5,1 | 3,5 | 1,6 |
| 4,1 | 4,2 | 3,0 | 1,1 |
‘ 1Ч>М',М''-гексаметнлпронзводное парафукснна (10.5).
зывают на то, что местом действия аминоакридинов является внешняя поверхность плазматической мембраны бактерии. Во- первых, антибактериальная активность этих соединений не снижается при повышении степени ионизации от 70 до 100%, несмотря на то, что при этом исчезают нейтральные молекулы, способные легче катиона проникать через мембрану. А во-вторых, при увеличении липофильности аминоакридинов происходит резкое уменьшение их антибактериального действия [Albert et al., 1945].
В бактериях большая часть ДНК клетки содержится в единственной кольцеобразной хромосоме, присоединенной к цитоплазматической мембране в одной точке [Ryter, Jacob, 1964]. РНК содержится в цитоплазматической мембране [Yudkin, Davis, 1965] и в рибосомах.
Профлавин (30 мкМ) in vitro ингибирует бактериальную ДНК-полимеразу на 85%, а РНК-полимеразу — на 30% [Hurwitz et al., 1962]. Некоторые наиболее типичные данные для РНК- систем представлены на рис. 10.4. На вставке показан график зависимости изменения величины обратной скорости полимеризации от обратной концентрации ДНК-матрицы, указывающий на то, что именно матрица является местом действия профлавина, ингибирующего таким образом синтез ДНК и РНК в живых бактериях. Механизм интеркаляции, с помощью которого аминоакридины соединяются со спиралью ДНК и препятствуют разделению ее цепей, будет рассмотрен в разд. 10.3.2.
Более подробно был изучен механизм действия аминоакридинов на ДНК-зависимый синтез РНК на примере Е. coli [Nicholson, Peacocke 1966], бактериофага [Sarris, Niles, Canellakis, 1977], Azotobacter [Canellakis et al., 1976]. Хотя ингибирование ферментов у эукариот нетипично, тем не менее 9-аминоак- ридин (500 мкМ) полностью ингибирует включение уридинмоно- фосфата (УМФ) при попытках проведения транскрипции ДНК вилочковой железы теленка РНК-полимеразой печени крысы [Zoncheddu et al., 1980].
Рассмотрим вкратце некоторые выдвигавшиеся ранее гипотезы о механизме действия аминоакридинов (помимо идеи об S6
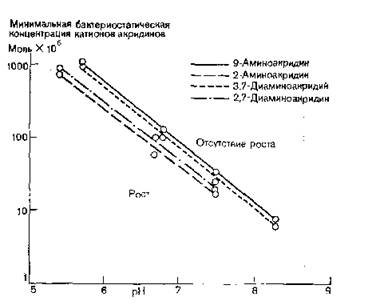
Рис. 10.3. Конкуренция между ионами водорода и катионами акридинов (организм Е. coli).
интеркаляции этих соединений в ДНК), представляющие интерес хотя бы с исторической точки зрения. Предполагалось, что антибактериальное действие акридинов обусловлено их способностью ингибировать различные ферменты. Однако для этого обычно требуются очень высокие концентрации аминоакридинов, в то время как даже в низких концентрациях эти соединения подавляют синтез и нормальное функционирование нуклеиновых кислот. [Albert, 1966].
Были сделаны попытки связать антибактериальное действие аминоакридинов с их окислительно-восстановительными потенциалами [Breyer, Buchanan, Duewell, 1944].
Так, для веществ с сильным антибактериальным действием (например, 3-амино- 9-амино- и 3,6-диаминоакридин) Ео' на первой стадии восстановления при pH 7,3 лежит между —0,47 и —0,92 В, тогда как соответствующее значение Ео' для соединений со слабым антибактериальным действием— между —0,31 и —0,39 В[4]. Таким образом, очень небольшое различие окислительно-восстановительных потенциалов (между —0,47 и —0,39 В) должно определить, обладает акридин антибактериальным действием или нет. Однако из рис. 10.3 видно, что в действительности не это различие определяет их активность. Так, при pH 7,3 одни акридины (например, 2-аминоакридин), слабо ионизированные при данном значении pH, обладают слабым, а другие — сильным антибактериальным действием, но активность их становится одинаковой при понижении pH до такой величины (например, 5,4), при которой обе группы акридинов сильно ионизированы. Поскольку снижение pH не уменьшает разности окислительно-восстановительных потенциалов 2- и 3-аминоакридинов,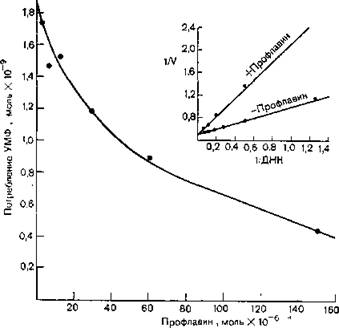
Рнс. 10.4. Ингибирование РНК-полимеразы Е. coli профлавином, измеренное по уменьшению потребления УМФ в присутствии избытка ДНК, АТФ, ГДФ и ЦТФ. На вставке представлен график зависимости обратной скорости полимеризации от обратной концентрации ДНК-матрицы в присутствии (верхняя линия) и в отсутствие (нижняя линия) профлавина [Hurwitz et al., 19621.
следует заключить, что окислительно-восстановительные потенциалы не влияют на антибактериальную активность производных акридина.
Гипотеза о том, что антибактериальное действие аминоакридинов определяется наличием свободных радикалов, образующихся на первой стадии (1Н) восстановления, также неверна, так как на этой стадии все акридины образуют свободные радикалы [Кауе, 1950]. Более того, вещества, не поддающиеся восстановлению, такие как 2-антранилгуанидин (табл. 10.10), проявляют акридиноподобное антибактериальное действие. Примечательно, что антрацены не восстанавливаются даже при —2,0 В [Zanker, Schnith, 1959].
Еще по теме Б. Ингибирование ДНК- и РНК-полимеразы.:
- 13. Переосаждение ДНК (РНК).
- Как избежать контаминации и деградации РНК и ДНК
- 3. Выделение ДНК и РНК из клеточных культур.
- 11.4. Гибридизация ДНК (РНК), иммобилизованной на мембране.
- Ингибиторы синтеза белка, не образующие аддукты ДНК и РНК
- Ингибиторы синтеза белка и клеточного деления, образующие аддукты ДНК и РНК.
- Механизмы ингибирования синтеза дигидрофолиевой кислоты сульфаниламидными препаратами
- Синтез кДНК на матрице суммарной РНК (задача 2)
- Методические рекомендации по постановке тестов ингибирования роста бактерий, выделенных в ветеринарных лабораториях при диагностике болезней животных
- Строение и функции РНК
- Ферментативный гидролиз РНК
- Выделение РНК (задача 1)
- Роль микро-РНК в патогенезе болезни Паркинсона
- Оценка качества суммарной РНК (задача 1)
- Технологии получения дрожжевой РНК