Б. Механизм связывания акридинов и их аналогов с нуклеиӯновыми кислотами.
В экспериментах in vitro было продемонстӯрировано, что связывание профлавина (10.7) (3,6-диаминоакриӯдин) с ДНК идет по двум механизмам: во-первых, по реакции первого порядка, в которой равновесие достигается при соотноӯшении 4 или 5 нуклеотидов на одну молекулу профлавина, и, во-вторых, в результате более медленной реакции высокого порядка, в которой одна молекула профлавина связывается с одной молекулой нуклеотида [Peacocke, Skerrett, 1956].
Второй процесс заключается в неупорядоченной адсорбции дополниӯтельных молекул акридина на внешней стороне спирали ДНК.
2-Антрилгуанидин
(10.21)
Этндий
(10.23)
В 1961 г. Lerman предположил, что молекулы 3,6-аминоак- ридина более прочно присоединяются к ДНК, благодаря интерӯкаляции между двумя слоями пар азотистых оснований, при этом первичные аминогруппы связываются ионной связью с двумя остатками фосфорной кислоты в спирали Уотсона — Криӯка, а плоский скелет акридинового цикла удерживается на моӯлекулах пурина и пиримидина ван-дер-ваальсовыми силами [Lerman, 1964а]. На рис. 10.7 представлена эта структура (вид сбоку). Необходимость большой плоскости поверхности и высоӯкой степени ионизации у молекул, обладающих антибактериӯальным действием, очевидно, и объясняется образованием этой структуры.
По данным рентгеноструктурного анализа слои азотистых оснований ДНК обычно связаны между собой сверху и снизу
Рис. 10.7. Схема вторичной структуры норӯмальной ДНК (слева) и ДНК, включающей интеркалироваиные молекулы профлавина (справа). Спираль рассматриваетси с отдаленӯной точки под таким углом, что видны лишь боковые проекции пар азотистых оснований и интеркалированиых молекул профлавина, а фосфорибозильный остов выглидит как регуӯлярная спираль.
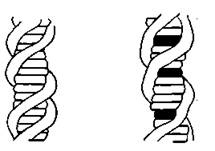
ван-дер-ваальсовыми силами. Расстояние между центрами атоӯмов соседних пар оснований в цепи составляет 0,336 нм. Это означает, что молекулы аминоакридинов, имеющие точно такую же толщину, что и пуриновые и пиримидиновые основания, могут проникать в оставшиеся 0,336 нм (от общего расстояния 0,672 нм). Это пространство может образоваться за счет неӯбольшого раскручивания двойной спирали (угол поворота варьируется для каждого вещества; так для этидиума он соӯставляет 26° [Wang, 1974], для профлавина и алкалоида зллип- тицина он несколько меньше [Kohn et al., 1975]).
Теперь рассмотрим, как Lerman пришел к теории интеркаӯляции. При взаимодействии профлавина с ДНК происходит трехкратное увеличение вязкости. Это объясняли тем, что внедӯряющиеся молекулы не только вытягивают спираль, но и делаӯют ее жесткой и спрямленной. Было установлено, что комплекс ДНК —профлавин имеет более низкий коэффициент седименӯтации, чем свободная ДНК- Это происходит за счет потери массы на единицу длины (ОММ профлавина составляет менее половины массы равного объема ДНК). Эти результаты были получены в разбавленном водном растворе. Было также обнаӯружено, что нити, вытягиваемые из комплекса, дают значительӯно более простые рентгенограммы, чем получаемые для чистой ДНК. Меридианальный рефлекс, соответствующий расстоянию 0,34 нм между соседними слоями, сохраняется, но новые полоӯжения первых экваториальных рефлексов свидетельствуют о том, что каждая молекула ДНК теперь имеет более плотную упаковку, а следовательно, и меньший диаметр, чем молекула чистой ДНК [Lerman, 1961]. В 1963 г. Lerman показал, что соотношение интенсивностей флуоресценции движущегося и неподвижного раствора согласуется с представлением о перпенӯдикулярном расположении молекул акридина по отношению к оси спирали. Затем он обнаружил резкое падение скорости диазотирования первичных аминогрупп профлавина в присутстӯвии ДНК- Это означало, что ДНК защищает аминогруппы от действия азотистой кислоты [Lerman, 1964].
Было замечено, что для денатурации ДНК после образования комплекса с 9-аминоӯакридином требуется более высокая температура [Lerman, 1964b] (ср. Chambron, ниже).Теория интеркаляции была подтверждена данными, полученӯными другими методами. Так, радиоавтография ДНК (содерӯжащих [3Н]тимин), выделенной из Т2-колифага, проводилась до и после погружения в разбавленный раствор профлавина. При этом было показано, что аминоакридин удлиняет молекулу ДНК от 45 до 75 мкм [Cairns, 1962]. Столь же убедителен был и тот факт, что благодаря интеркаляции профлавина темпераӯтура плавления (Тт) ДНК повышалась на 20°, а при «расплавӯлении» комплекса происходило внезапное выделение большей части связанного профлавина (т. е. нити двойной спирали разӯделялись) [Chambron, Daune, Sadron, 1966]. Впоследствии это явление было подтверждено Kleinwachter, Bakarova, Bohacek (1969).
Прямое доказательство правильности теории интеркаляции было получено при рентгеноструктурном изучении комплекса профлавин — ДНК., показавшем, что молекула аминоакридина располагается параллельно парам оснований ДНК в соотноӯшении 1:3 [Neville, Davies, 1966]. Исследование растворов комплексов пяти моноаминоакридинов с ДНК методами линейӯного и кругового дихроизма подтвердило, что катионы акридиӯна лежат в плоскостях, параллельных тем, в которых находятся пары оснований [Jackson, Mason, 1971]. И наконец, было обнаӯружено, что свободная энергия связывания аминоакридинов с ДНК близка энергии, характерной для процессов интеркаляӯции, но слишком высока для любого другого вида связывания на внешней стороне цепей ДНК [Jordan, 1968].
С точки зрения электростатических взаимодействий область ДНК, доступная для интеркаляции, несет высокий отрицательӯный заряд, образующийся не только за счет фосфат-анионов, пуриновых и пиримидиновых оснований (за исключением адеӯнина) и частичных отрицательных зарядов атомов кислорода дезоксирибозы. Именно в это отрицательно заряженное простӯранство и втягиваются катионы лекарственного вещества, почӯти полностью нейтрализуя избыточный отрицательный заряд.
Последующее образование ван-дер-ваальсовых связей с основаӯниями ограничивает дальнейшее продвижение молекулы и обесӯпечивает относительную устойчивость комплекса. На рис. 10.8 показана интеркаляции профлавина в молекулу ДНК (изобраӯжение построено по данным рентгеноструктурного анализа). Согласно расчетам, средняя площадь пары оснований в спирали ДНК или РНК составляет около 0,5 нм2, что превышает плоӯщадь молекулы акридина (0,385 нм2), рассчитанную метоӯдом, показанным на рис. 10.5. Однако в любой одноцепочечной модели небольшая часть каждой молекулы акридина будет выӯпячиваться наружу. Липофильные аналоги, такие как акридиӯновый оранжевый (3,6-бисдиметиламиноакридин), не только интеркалируют, но и прочно прикрепляются к внешней стороне ДНК.В. Другие соединения, интеркалирующие в ДНК. Как это часто случается с новыми идеями, концепция интеркаляция поӯначалу не получила должного признания. Однако вскоре, и это
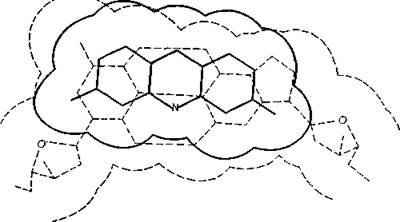
Рис. 10.8. Интеркаляции 3,6-диамииоакридииа между слоями пар оснований ДНК. Левое кольцо акридина противостоит почти точно иад цитозином, а праӯвое кольцо — почти точно иад пиримидиновым циклом гуанина; область ван- дер-ваальсовых связей молекулы акридина обозначена сплошной линией [Ler- тап, 1964Ь].
тоже не редкость, после ее признания очень многим лекарственӯным веществам самых разных типов стали приписывать именно этот механизм действия. Для проверки подобных утверждений был введен специальный тест на интеркаляцию, показывающий, вызывает ли исследуемое вещество локальное раскручивание двойной спирали суперспирализованной кольцевой ДНК, наприӯмер ДНК колифага [Waring, 1970]. При первичном добавлении аминоакридина количество правозакрученных суперспиралей равномерно уменьшается; при достижении критического соотноӯшения добавленных молекул суперспирали исчезают и ДНК превращается в раскрученное открытое кольцо, имеющее благоӯдаря интеркаляции большой диаметр.
По мере дальнейшего добавления лекарственного вещества нарастание напряжений в кольце ДНК снова приводит к появлению суперспиралей, но теперь уже левозакрученных. Так как суперспирали более комӯпактны, чем раскрытые кольца, они быстрее осаждаются, что позволяет судить о ходе процесса по изменению коэффициента седиментации, проходящего через минимум [Waring, 1970].Применение этого метода позволило установить, что мехаӯнизм интеркаляции характерен для некоторых аминофенантри- динов, включая (10.23), для антималярийного препарата хин- гамин (10.31), трех карциностатических антибиотиков: ногало- мицина, даунорубицина и актиномицина D (4.38) (разд. 4.0). Однако для хлорпромазина и диэтиламида лизергиновой кислоӯты (ЛСД) такого подтверждения получено не было, хотя ранее им приписывали интеркаляционный механизм действия. Было обнаружено, что спермин, стрептомицин, диминазен и митрами- цин активно взаимодействуют с ДНК, но не образуют интерка- лятов [Waring, 1970].
Рассмотрим другие, не менее важные примеры интеркаля- ции. Парафуксин (10.5), краситель трифенилметанового ряда, интеркалирует в ДНК, однако не так плотно, как профлавин [Armstrong, Panzer, 1972]. Антибиотик эхиномицин содержит полипептидную цепь, с которой связаны два хиноксалиновых цикла, далеко отстоящие друг от друга. Эти циклы одновреӯменно интеркалируют в разные участки ДНК. Происходящее вследствие этого раскручивание и удлинение спирали в два раза превышает то, которое вызывается действием аминоакриӯдинов [Waring, Wakelin, 1974].
Путем введения акридиновых циклов в молекулы путресци- на (11.6) и спермина (11.4) были получены синтетические анаӯлоги, способные ингибировать ДНК-зависимую РНК-полимераӯзу и повышать температуру плавления ДНК сильнее, чем сами аминоакридины [Canellakis et al., 1976]. Противораковое дейстӯвие соединений, образующих двойные интеркаляты, рассматӯривается в разд. 10.3.4.
Биологические свойства лекарственных веществ, действие которых основано на интеркаляции, обсуждаются в обзоре [Schwartz, 1979].
Почему же аминоакридины и соединения с подобным мехаӯнизмом действия обладают такой высокой избирательностью действия по отношению к бактериям и практически не дейстӯвуют на клетки млекопитающих? Установлено, что для серии аминоакридинов характерно наличие антибактериальных свойств, высокая степень ионизации и интеркаляционный мехаӯнизм действия [Jackson, Mason, 1971]. Избирательность их дейӯствия обусловлена, по-видимому, двумя факторами: доступӯностью одиночных хромосом бактерий (разд. 5.3), а также тем, что эти лекарственные вещества действуют преимущественно на кольцевые ДНК [Waring, 1970]. Кроме избирательного дейӯствия на бактерии, аминоакридины действуют избирательно и на другие кольцевые ДНК: они способны отщеплять бактериӯальные плазмиды [Bouanchaud et al., 1968], а также вызывать наследственные изменения у дрожжей путем подавления реплиӯкации ДНК в митохондриях, не затрагивая при этом ядра (разд. 10.3.1) [Hollenberg, Borst, van Bruggen, 1970].
Далее, аминоакридины и аминофенантридины могут или поӯдавлять кинетопласт в трипаносомах или (при более низких концентрациях) разрушать его кольцевую ДНК [Riou, Delain, 1969].
Рассчитано, что, поскольку эти лекарственные вещества уменьшают суперспирализацию кольцевой ДНК, выделение свободной энергии повышает сродство к агенту, в то время как аналогичная однонитевая ДНК имеет пониженное сродство [Bauer, Vinograd, 1970]. По-видимому, этот случай можно отӯнести к проявлению избирательности за счет накопления (гл. 3).
10.3.3.
Еще по теме Б. Механизм связывания акридинов и их аналогов с нуклеиӯновыми кислотами.:
- Механизмы ингибирования синтеза дигидрофолиевой кислоты сульфаниламидными препаратами
- Катионные антибактериальные средства, действующие ло типу акридинов.
- Нарушения обмена гематоидина, гемати- нов и гематопорфирина
- Реакция связывания комплемента
- Г. Применение акридинов в современной медицине.
- Количественные аспекты связывания металлов
- Нейропротекторное действие нейролипинов и их аналогов
- Нейрозащитное действие простамида и его аналогов
- Аналоги витаминов и коферментов
- Аналоги аминокислот
- 9.4.5. Аналоги пуринов и пиримидинов
- Аналоги гормонов и нейромедиаторов
- 9.4.7. Другие аналоги
- Влияние субхронического системного введения пептида TGeNHR-NH2 на характеристики рецепторного связывания на мембранах мозга мышей линий C57Bl/6 и Balb/c методом ex vivo анализа
- Аналоги гонатропин-ризилинг гормона