Роль фолиевой кислоты и других птеридинов
Как было показано выше, производные фолиевой кислоты играют ключевую роль в биосинтезе пуринов и пирймидинов. Эти птеридины представляют собой коферменты, ответственные за включение атомов углерода в положения 2 и 8 в молекулах пуринов и за введение метильной группы в молекулу уридина при синтезе тимидина (4.1,6).
При действии на бактерии суль- 32фаниламидных препаратов в низких концентрациях в культуральной жидкости накапливается рибонуклеотид 4-аминоими- дазол-5-карбоксамида (9.22). Это соединение является полупродуктом биосинтеза инозиновой кислоты (9.23), из которой образуются все пурины.
Коферменты образуются из дигидрофолиевой кислоты (9.20), которая превращается в тетрагидрофолиевую кислоту под действием фермента дигидрофолатредуктазы [Osborn, Freeman, Huennekens, 1958]. Тетрагидрофолиевая кислота затем модифицируется различными заместителями, содержащими один атом углерода. Роль кофермента, обеспечивающего введение атома С-2 в пурины, т. е. формилирования риботила (9.22) с образованием инозиновой кислоты (9.23), играет №°-формил-5,6,7,8- тетрагидрофолиевая кислота. На более ранней стадии введения атома С-8, т. е. формирования риботида, в результате которого (в конечном счете) образуется риботид соединения (9.22), участвует тот же фермент, а не Ы5,Н10-метенилтетрагидрофолиевая кислота (9.24), как полагали ранее. Коферментом, обеспечивающим введение метильной группы в молекулу уридиловой кислоты, т. е. образование тимидиловой кислоты, является N5,N10-Me- тилентетрагидрофолиевая кислота (9.25), которая ответственна также за взаимопревращения двух аминокислот — серина и глицина. Метилирование уридилата с образованием тимидила- та происходит под действием тимидилатсинтетазы (ОММ 67 ООО). Синтез метионина и катаболизм гистидина также катализируются птеридиновыми коферментами, каждый из которых действует совместно со специфичным апоферментом.
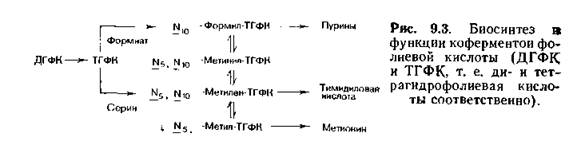
Фолиевые КИСЛОТЫ, содержащие полипептиды, образованные из остатков глутаминовой кислоты, по-видимому, выполняют еще не выясненные функции коферментов. В плазме человека дигидрофолие- вая кислота содержит лишь один остаток глутаминовой кислоты, однако в природе число этих остатков часто увеличивается до 3—7; особенно это характерно для бактерий [Kisliuk, 1981]. Основное количество фолиевой кислоты человек получает с овощами в виде полиглутамилфолиевой кислоты. Богатыми источниками служат также печень и дрожжи. Биосинтез и функции фолиевых коферментов представлены на рис. 9.3.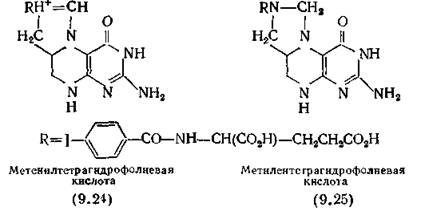
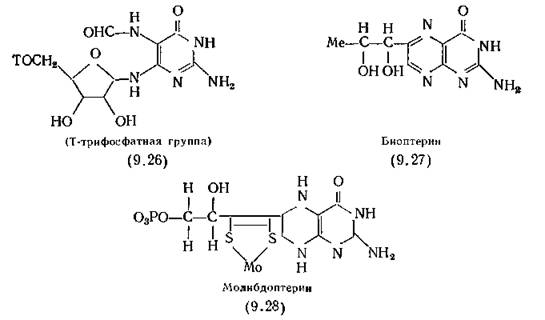
В природе биологически активные птеридины образуются при деградации гуанина (4.4) (в виде GTP), приводящей к получению 2-амино-4-оксо-6-(5-О-трифосфо-Е)-рибозиламино)-5~ формиламинопиримидина (9.26) [Shiota et al., 1969b). Под действием фермента дигидронеоптеринтрифосфатсинтетазы (получаемой из Lactobacillus plantarum) он циклизуется за счет образования связи между экзоциклическим атомом азота и атомом С2 рибозы. Синтезированный таким образом дигидронеоп- терин 2-амино-7,8-дигидро-4-оксо-6-(П-эритро-1',2',3'-тригидрок- сипропил)-птеридин под действием фермента гидронеоптериналь- долазы (получаемой из бактерий Е. coli) выделяет гликолевый альдегид, а остаток 2-амино-4-оксо-6-гидроксиметил-7,8-дигидро- птеридин легко встраивается в молекулу фолиевой кислоты (разд. 9.3.1).
Участие птеридинов в метаболизме не ограничивается производными фолиевой кислоты. Следует отметить, что наличие аминогруппы в положении 2 гетероцикла не имеет особого значения для проявления биологической активности. Так, биосинтез витамина рибофлавина осуществляется через 6,7-диметил-2,4- дезокси-8-рибитилптеридин.
В этой реакции одна молекула пте- ридина предоставляет, а другая принимает четыре атома углерода для образования орто-ксилольного цикла рибофлавина [Plaut, 1964]. Исходным соединением для данного птеридина служит GTP, рибозильная часть которого превращается в ри- битил-группу молекулы рибофлавина.Биоптерин (9.27)— птеридин цвета слоновой кости, образуется в процессе биосинтеза у животных и широко распространен в природе; его химическое название—Ь-эритро-2-амино- 6-(1,2-дигидроксипропил)-4-оксоптеридин. Исходным веществом для синтеза этого птеридина также является GTP. Восстановленная форма 5,6,7,8-тетрагидробиоптерина служит кофактором, необходимым для ряда ферментов, использующих молекулярный кислород для окисления фенилаланина в тирозин [Kaufman, 1964], тирозина в дофаминкарбоновую кислоту (3.43,6), триптофана в 5-гидрокситриптофан, а также, возможно, и при синтезе меланина. Этот кофактор использует молекулярный кислород. Его применяют в клинике при лечении одного из видов кетону- рии у детей [Cederbaum, 1979]. Тетрагидробиоптерин улучшает состояние больных при паркинсонизме [Narabayashi, 1982].
Птеридин считают основным рецептором электронов, высвобождающихся под действием света в процессе фотосинтеза [Fuller et al., 1971]; предполагают также, что птеридин, содержащийся в глазах млекопитающих, защищает их от ослепляющего действия света [Cremer-Bartels, 1975]. Раковые клетки расщепляют биоптерин до неоптерина, концентрация которого в моче может служить показателем при диагностике рака [Rokos et al., 1980].
Другим важным природным птеридином является молибдо- птерин (9.28), кофермент ксантиндегидрогеназы, альдегидокси- лазы, нитратредуктазы, сульфитоксидазы и, видимо, других ферментов, для действия которых необходимо наличие молибдена и железа [Johnson, Rajagopolan, 1982]. Ксантоптерин — колим- фокин (разд. 5.2), ингибирует размножение лимфоцитов [Ziegler et al., 1983].
Химия и биология птеридинов см. Blair (1983).
Химия птеридинов во многих отношениях необычна, главным образом из-за наличия в их молекулах четырех двоесвязанных атомов азота, обладающих сильно выраженными электроноакцепторными свойствами, вследствие чего нарушается ароматический характер этих соединений.
Для птеридинового кольца характерна тенденция к ковалентному присоединению молекулы воды по двойной связи даже при комнатной температуре, например, ксантоптерин (разд. 2.5.1) [Albert, 1967, 1976]. Несколько легче происходит присоединение таких веществ, как ацетон, кетокислоты и меркаптаны. Еще одна особенность птеридинов и некоторых других азотсодержащих гетероциклов заключается в понижении их растворимости в воде за счет эффектов заместителей, способных к образованию внутри- или межмолекулярных водородных связей (например, —NH2 или —NH—С = О группы) [Albert, Brown, Cheeseman, 1952]. У природных птеридинов обнаружена
и количественно охарактеризована выраженная способность к хелатообразованию [Albert, 1953].
Биохимия фолиевой кислоты и родственных ей птеридинов см. Blakley (1969).
9.3.3.
Еще по теме Роль фолиевой кислоты и других птеридинов:
- Применение фолиевой кислоты
- 9.3. Антагонисты фолиевой кислоты
- Роль цитокинов и других гуморальных факторов в развитии кахексии.
- ГЛАВА XIV. РОЛЬ КОРЫ БОЛЬШИХ ПОЛУШАРИЙ И ДРУГИХ ОТДЕЛОВ МОЗГА В ОБРАЗНОЙ ПРОСТРАНСТВЕННОЙ ОРИЕНТАЦИИ
- РОЛЬ ОПУХОЛЕВЫХ МАРКЕРОВ И ДРУГИХ КЛИНИКО-ЛАБОРАТОРНЫХ ПОКАЗАТЕЛЕЙ В ПРОГНОЗИРОВАНИИ РЕЦИДИВИРОВАНИЯ ПЛОСКОКЛЕТОЧНОГО РАКА ГОЛОВЫ И ШЕИ
- Название нозологической единицы: Наружный отит при других болезнях, в других рубриках. Н-62.4
- Название нозологической единицы: Средний отит при других болезнях, классифицированных в других рубриках. Н-67.8
- Название нозологической единицы: Наружный отит при других инфекционных и паразитарных болезнях, классифицированных в других рубриках. Н-62.3
- Механизмы ингибирования синтеза дигидрофолиевой кислоты сульфаниламидными препаратами
- Жирные кислоты