Влияние субхронического системного введения пептида TGeNHR-NH2 на характеристики рецепторного связывания на мембранах мозга мышей линий C57Bl/6 и Balb/c методом ex vivo анализа
Выбор лиганда для радиорецепторных исследований ex vivo осуществляли в соответствии с рекомендациями IUPHAR. При радиолигандном анализе NMDA- рецепторов использовали [3Н^](+)МК-801, синтезированный реакцией ВТКИО с молярной радиоактивностью 210 Ки/ммоль (Золотарев и др., 2009).
Молярная радиоактивность этого препарата на порядок выше, чем у коммерчески доступного селективно меченного тритием препарата фирмы NEN. Для анализа серотонино- вых 5-HT2a и никотиновых н-АХ-рецепторов использовали ^-3Н]кетансерин и (-) ^-3Н]никотин с молярной радиоактивностью 114 и 140 Ки/ммоль, соответственно (Zolotarev et al., 2010).Рецепторный компонент механизма действия пептида был проведен сравнительным анализом изменения плотности NMDA-глутаматных, 5-HT2a-серотонино- вых и н-АХ-никотиновых рецепторов на мембранах префронтальной коры и гиппокампа мозга мышей инбредных линий C57Bl/6 и Balb/c. Мыши линии Balb/c с исходным когнитивным дефицитом характеризуются меньшей плотностью исследуемых рецепторов.
Плотность мест связывания [G-3H]MK-801 с NMDA-рецепторами в гиппокампе мышей линии Balb/c меньше на 23%, чем у мышей линии C57Bl/6. Плотность мест связывания 5-HT2a-серотониновых и н-АХ-никотиновых рецепторов на мембранах префронтальной коры мозга также оказалась на 30-35% меньшей у мышей линий Balb/c, чем у мышей линии C57Bl/6.
Ткань гиппокампа гомогенизировали в гомогенизаторе Potter S («тефлон-стекло») в 10 объемах 5 мМ HEPES и 4,5 мМ Tris-base (рН=7,6), содержащего 0,32 М сахарозу (буфер №1). Гомогенат разбавляли до 50 объемов буфера для исследования 2 (5 мМ HEPES и 4,5 мМ Tris-base) (рН=7,6) и центрифугировали 10 мин при 1000 g. Супернатант отбирали и вновь центрифугировали 20 мин при 25 000 g. Осадок гомогенизировали в 50 объемах буфера 2 и центрифугировали 20 мин при 8 000 g.
Супернатант и его мягкий, зыбкий надслой отбирали и центрифугировали 20 мин при 25 000 g. Полученный осадок суспендировали в буфере 3 (5 мМ HEPES и 4,5 мМ Tris-base, содержащем 1 мМ Na4EDTA, рН=7,6 и суспензию вновь центрифугировали. Такую процедуру отмывки проводили четырежды, причем при последней отмывке EDTA был исключен из состава. Конечный осадок ресуспенди- ровали в 5 объемах буфера 2 и сохраняли в жидком азоте.Реакционная смесь содержала 200 мкл буфера 2, 50 мкл меченого лиганда (50 нМ) и 250 мкл белковой суспензии. Неспецифическое связывание определяли в присутствии 50 мкл немеченого лиганда. Инкубационная смесь содержала 200 мкл буфера 2, 50 мкл меченого лиганда (50 нМ) и 250 мкл суспензии белка. Неспецифическое связывание определяли в присутствии 50 мкл немеченого (+)МК-801. Специфическое связывание рассчитывали как разницу между общим и неспецифическим связыванием. Измерение радиоактивности осуществляли жидкостно-сцинтилляцион- ным методом на счетчике TriCarb TR290. Эффективность счета составляла 42-47%. Показатели количества мест связывания лиганда (Bmax, фмоль/мг белка) и константу диссоциации лиганд-рецепторных комплексов (Kd, нМ) рассчитывали с помощью программы GraphPadPrizm 4 Demo, статистическую обработку проводили по программе Statsoft 6,0. Концентрацию белка определяли стандартным методом Лоури.
Результаты радиорецепторного анализа связывания лиганда [3Н^](+)МК801с NMDA-рецепторами, участвующими в регуляции когнитивных форм поведения, были представлены после обработки по Скетчарду (рис. 9).
Анализ влияния на NMDA-рецепторы гиппокампа мышей линии Balb/c показывает, что субхроническое введение пептида приводит к возрастанию величины
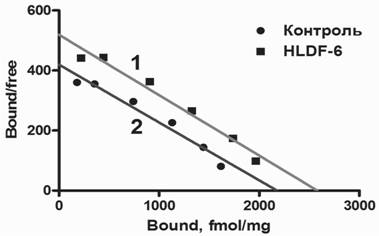
Рис. 9. График Скетчарда, отражающий влияние субхронического внутрибрюшинного введения пептида TGeNHR-NH2 на характеристики рецепторного связывания [3H-G](+)MK-801 с NMDA-рецепторами на мембранах мозга гиппокампа мышей линии Balb/c
Bmax, отражающей плотность соответствующих рецепторов, с 2115±36 фмоль/мг белка в контрольной группе до 2527±43 фмоль/мг белка после субхронического введения препарата.
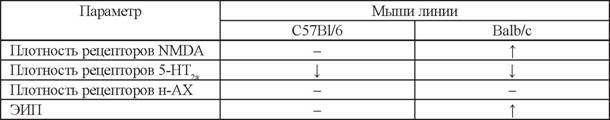
В то же время, в мозге мышей С57В1/6 изменения в рецепторных характеристиках не отмечаются: плотность мест связывания лиганда в контрольной группе - 2734±44 фмоль/мг, а после субхронического введения препарата эта величина составила 2625±55 фмоль/мг белка.После 5-кратного системного введения амидной формы пептида TGeNHR-NH2 в ежедневной дозе 100 мкг/кг величина плотности мест связывания [G-3H]MK-801 с NMDA-рецепторами в гиппокампе мышей Balb/c увеличилась на 21% относительно интактного контроля. При этом величина плотности NMDA-рецепторов в гиппокампе мышей Balb/c приблизилась к соответствующему значению для группы сравнения, представленной мышами линии C57B1/6, относительно которой препарат был неэффективен. Субхроническое введение пептида в равной степени уменьшало плотность 5-НТ^-серотониновых рецепторов у обеих линий и не оказывало влияния на никотиновые н-АХ-рецепторы (табл. 1). Результаты, полученные при исследовании влияния амидной формы пептида HLDF-6 на нейрохимические процессы с помощью радиолигандного анализа, показывают, что в механизм этих фармакологических эффектов с высокой степенью вероятности вовлечена глута-
Таблица 1
Влияние субхронического системного введения пептида TGeNHR-NH2 на изменение плотности глутаматных (NMDA), серотониновых (5-HT2a) и никотиновых рецепторов
(н-АХ) на мембранах префронтальной коры и гиппокампа мозга, а также эффективность исследовательского поведения (ЭИП) мышей инбредных линий C57Bl/6 и Balb/c
матергическая система гиппокампа, в частности NMDA-тип рецепторов. NMDA- рецепторы являются одной из важнейших мишеней для воздействия ноотропных и нейропротекторных лекарственных препаратов, так как они принимают определяющее участие в регуляции уровня выживаемости и гибели
6.
Еще по теме Влияние субхронического системного введения пептида TGeNHR-NH2 на характеристики рецепторного связывания на мембранах мозга мышей линий C57Bl/6 и Balb/c методом ex vivo анализа:
- Изучение влияния субхронического системного введения пептидов TGENHr-NH2, ENHR-NH2 и HR-NH2 на эффективность исследовательского поведения мышей линий C57Bl/6 и Balb/c
- Поиск первичной мишени пептида TGeNHR-NH2 и ex vivo анализ изменения рецепторных характеристик мозга при его введении
- Сравнительное изучение времени жизни в плазме крови крысы пептидов TGENHR и TGeNHR-NH2
- Влияние нетрадиционных методов введения химиопрепаратов на синтетическую активность клеток крови и костного мозга интактных крыс
- Влияние нетрадиционных методов введения нейпогена . на клеточный состав костного мозга животных без опухоли
- Влияние острого введения алкоголя на личностные свойства и системную организацию поведения в реальной и виртуальной средах
- Математический метод извлечения терминалей аксонов ДА-ергических нейронов в области стриатума срезов головного мозга мышей
- Исследование фармакокинетики и фармакодинамики амидной формы нейропротекторного пептида HLDF-6 с использованием равномерно меченных изотопами водорода соединений
- Влияние острого введения алкоголя на системные процессы, обеспечивающие выполнение задачи «сенсомоторного выбора»
- 3.3.3. Опыты с субхроническим введением веществ
- 3.3.4. Опыты с субхроническим сочетанным введением веществ
- Опыты с субхроническим сочетанным введением антагониста NMDA- рецепторов нерамексана и морфина
- Опыты с субхроническим введением антагонистов NMDA-регіепторного комплекса (мемантин, нерамексан)
- НЕЙРОТРОФИЧЕСКИЙ ПЕПТИД ИЗ ПОВРЕЖДЕННОГО МОЗГА
- 3.5. Однофакторный дисперсионный анализ хронического влияния барбитуратов на ядра основных отделов миндалевидного тела головного мозга крыс в различные возрастные периоды
- Влияние острого введения алкоголя на личностные свойства, выявляемые с помощью психодиагностических методов
- Опыты с однократным введением антагонистов NMDA-рецепторного комплекса
- Характеристика токсичности и биологического действия фуллеренов in vivo
- Влияние ГУМТ на концентрацию глюкозы и С-пептида в пуповинной крови