Естественная и антропогенная трансформация Прикаспийского песчаного очага чумы.
Формирование нынешнего облика обоих очагов чумы СевероЗападного Прикаспия происходило в верхнем голоцене (Дмитриев, 2000; 2001), так же, как и остальной территории Прикаспийской низменности.
Вопрос о времени приобретения самостоятельности песчаным очагом не вполне ясен и даже дискуссионен. Одной из правдоподобных версий является признание одновременности и параллельности возникновения и развития сусликового и песчаночьего очагов чумы, по меньшей мере, с позднего плейстоцена, причем там в то время, присутствовали большая и краснохвостая песчанки (Тропин, 1968; 1971; 1975; Дмитриев, 1983; 2000; 2001). Для целей профилактики чумы в этом регионе достаточно знания его современного ландшафтного и эпизоотологического облика, поэтому вопрос о палеоистории очага мы не рассматриваем. Однако на протяжении новейшей истории оба очага функционировали самостоятельно и в целом не синхронно (Подсвиров с соавт., 1995), несмотря на то, что наиболее обширные эпизоотические проявления чумы иногда могли охватывать их территории одновременно и без пространственных разрывов. Мнение о том, что антропогенная трансформация ландшафтов, регистрируемая в последние десятилетия, послужила причиной возникновения нового песчаного очага чумы, несколько гиперболизировано (Буркин с соавт., 1982; Тихомиров с соавт., 1989). По нашему мнению, происходящие изменения если выходят за рамки обычных (естественных) колебаний ландшафно-климатических условий, то весьма незначительно, но они (изменения), в конце концов, обратили внимание исследователей на иной статус рассматриваемой территории и заставили признать это официально.Изменения в характере эпизоотических проявлений чумы, произошедшие в последние десятилетия в Северо-Западном Прикаспии, и их несинхронность в разных частях региона (Подсвиров с соавт., 1995) привлекали и продолжают привлекать внимание исследователей.
102
Основными причинами этих изменений являются два мощных фактора.
Первый из них — современная антропогенная трансформация ландшафтов (орошение, распашка, перевыпас мелкого рогатого скота и т. д.), вызвавшая очень большие и, возможно, необратимые изменения первичных степных биоценотических комплексов в этом регионе (Варшавский с соавт., 1986; 1991; Денисов с соавт., 1986; Попов Н.В. с соавт., 1989; Шилов с соавт., 1989;). Усиление антропогенного воздействия совпало по времени с другим фактором — возрастанием аридности климата в регионе (Дятлов, 1986), явившимся прямым следствием регрессии Каспийского моря. Резкое отступление Каспия в 30-60-е гг. прошлого века, вследствие которого к началу 70-х годов береговая линия отодвинулась в восточном направлении на несколько десятков километров, сопровождалось существенным падением уровня грунтовых вод (Козлова, 1994; Козлова с соавт., 1994; 1998).Продуктивность пастбищ Прикаспийской низменности в этот период значительно снизилась. Длительный неумеренный выпас скота привел к коренным изменениям структуры растительного покрова, на месте злаковых и полынно-злаковых ассоциаций стали формироваться эбелековые сбои. На легких супесчаных и песчаных почвах широкое распространение получили массивы развеянных песков (Виноградов с соавт., 1985). В то же время ирригация и мелиорация Прикаспийской низменности коренным образом преобразовала ландшафтную и биоценотическую структуры целых районов в другую сторону. Увеличение влажности способствовало смене растительных ассоциаций, во многих местах возникли заросли рогоза, тростника и другой околоводной растительности. Оно способствовало расселению в полупустыню таких мезофильных видов грызунов, как водяная полевка, ондатра, серая крыса (Варшавский с соавт., 1989). По валам оросительных каналов далеко на восток проникли полевая и лесная мыши (Яковлев, 1996). На забурьяненных залежах, по валам обводнительных каналов
103 возникли условия для постоянной концентрации разных видов грызунов (в первую очередь, малых песчанок и мышевидных).
При всем этом нельзя не отметить, что в последние 60-70 лет на территории Прикаспийской низменности (особенно в ее северо-западной части) неоднократно имели место значительные колебания численности фоновых видов грызунов — малого суслика, полуденной и гребенщиковой песчанок.
Это естественный и закономерный процесс, однако, отмечено, что наиболее глубокие депрессии численности зверьков и значительные изменения размеров их поселений неизменно происходили под комплексным влиянием климатических и антропогенных факторов (Варшавский с соавт., 1991).За последние 30-40 лет в результате зарегулирования стока рек, перевыпаса мелкого рогатого скота, регрессии Каспия и связанной с этим общей аридизацией ландшафта, усыханием и осолонением озер, развитием солончаков, появлением новых массивов развеваемых песков, сократились площади и плотности поселений малого суслика. Развитие поливного земледелия привело к появлению густой сети оросительных каналов и водогонов, вдоль которых произрастают ассоциации рудеральной растительности, тростниковые заросли и бурьянники. Брошенные водогоны без проведения агротехнических мероприятий послужили идеальным условием для увеличения численности мышевидных грызунов, в первую очередь домовой мыши. Расширение площадей развеваемых и полузакрепленных песков привело к повышению численности песчанок, особенно полуденных. Аналогичные процессы смены доминирующих видов происходили и на других территориях (Лисицын с соавт., 1969; Бурделов, Поле, 1985).
Из 26 видов обитающих здесь грызунов, три являются основными носителями возбудителя — полуденная и гребенщиковая песчанки (М. meridianusи М. Iamariscinus) и малый суслик (S. pigmaeus).В отдельные годы в эпизоотический процесс могут вовлекаться домовые мыши (М.
104 musculus),серые хомячки (Cricetulus migratorius), общественные полевки (Microtus Socialis),емуранчики (Scirtopoda telum)и другие животные.
Вопрос о гостальности и биоценотической структуре восточной и юго-восточной части очага чумы Северо-Западного Прикаспия существует с момента установления здесь факта наличия энзоотии. Начиная с 1923 года, когда началось исследование полевых грызунов на зараженность чумой, большинство культур возбудителя получено от малых сусликов и их блох. Значительное количество штаммов выделялось также от песчанок и мышевидных грызунов на Черных землях (Нарын-Худук), в Приморском и Ильменно-Придельтовом районах (Суворов, 1924; Павленко, 1959; Шишкин, 1959).
Однако, в те годы численность малого суслика, чьи поселения перемежались с поселениями песчанок и мышевидных грызунов, была значительно выше, чем сейчас. Поэтому, при отлове грызунов в первую очередь обращали внимание на сусликов, считавшихся единственными виновниками чумной энзоотии.До 1987 г. основным носителем в тогда еще едином очаге СевероЗападного Прикаспия считался малый суслик. После возобновления эпизоотической активности юго-восточной части очага, случившегося осенью 1979 г., стало ясно, что эта территория характеризуется иной биоценотической структурой населяющих ее носителей и переносчиков возбудителя, поддерживающих природную очаговость чумы (Лабунец с соавт., 1981; Илюхин с соавт., 1983; 1984). Заметим, что в эпоху раннего голоцена на территории Северного Прикаспия доминирующее положение занимала желтая пеструшка, чувствительный к чумной инфекции зверек (Дмитриев, 1983). Возможно, она играла здесь основную роль в поддержании природной очаговости чумы.
Начиная с 1987 г., было официально признано существование Прикаспийского песчаного очага с малыми песчанками в качестве основных носителей. Однако ареалы полуденной и гребенщиковой песчанок в этом регионе совпадают не полностью. Полуденная песчанка в
105 северной части Волго-Донского водораздела распространена севернее и западнее гребенщиковой и имеет изолированный участок ареала на Ергенях. В Терско-Кумском междуречье гребенщиковая песчанка проникает далеко на юг и запад, то есть, распространена гораздо шире полуденной, встречающейся в районе нижнего течения р. Кумы и в приморской полосе (Бакеев, 1953; 1956; Павлов, Климченко, Лисицын с соавт., 1957; Павлов, 1959). Закономерно, что наиболее активная часть очага приурочена к зоне совместных поселений двух видов песчанок (Лавровский, 1964), дислоцированных на востоке Черных земель, в Приморском и Ильменно-придельтовом районах Волго-Кумского и на севере Терско-Кумского междуречий.
Численность песчанок в очаге оценивается как более стабильная, чем в Волго-Уральских песках (Павлов, 1961; 1962; 1965).
Лишь в отдельные особо неблагоприятные годы численность полуденных песчанок может резко снижаться, затем быстро восстанавливаться і В центральных частях очага плотность полуденной песчанки может повышаться до 20-30 особей на га и выше. Обычно же она составляет 5-10 зверьков.в среднем. Гребенщиковая песчанка незначительно уступает полуденной в численности, особенно в засушливых частях очага. Однако в Ильменно- придельтовом и Приморском районах, покрытых более развитой растительностью, она часто доминирует над полуденной песчанкой. Южнее Кумы, особенно в западной части Терско-Кумского междуречья, гребенщиковая песчанка более многочисленна. Плотность ее населения нередко достигает 10-20, а на отдельных участках — 40-60 и более зверьков на га.Численность малого суслика — третьего по значимости основного носителя возбудителя в Прикаспийском песчаном очаге чумы — в последние десятилетия очень низка. Основными причинами снижения численности сусликов в Северо-Западном Прикаспии явились истребительные работы против этих зверьков и распашка целинных земель
106 (Миронов, 1959 а). В юго-восточной же части региона (современный песчаный очаг) истребление сусликов почти не проводилось, а причиной снижения их численности послужили естественные и антропогенные преобразования ландшафта, о которых мы писали выше.
Определенное эпизоотологическое значение в очаге имеют серые хомячки, емуранчики и общественные полевки. В отдельные годы среди этих грызунов встречались зараженные чумой, что было связано с их резко возраставшей численностью. Такое явление наблюдается достаточно редко, но оно, очевидно, способно несколько активизировать эпизоотический процесс (Илюхин с соавт., 1984; Матросов, Кузнецов, Слудский с соавт., 2003; Матросов, Попов, Кузнецов, Слудский, 2003). На территории природных очагов чумы известны хронологически отслеженные свидетельства изменения эпизоотологического значения тех или иных видов грызунов в случае вымирания или снижения численности основных носителей чумы (Ралль, 1960 а; Фенюк, 1963; Тропин, 1968; 1971; Дмитриев, 1983; 2001).
Увеличение антропогенного воздействия на биоценозы Северо-Западного Прикаспия привело к опустыниванию зональных ландшафтов и изменению ареалов и структуры населения животных (Зонн, 1995). Лишь в последнее десятилетие здесь началось восстановление растительности (Шилова с соавт., 2000; 2004).Естественно, что первыми на это откликнулись более пластичные мелкие полиэстральные виды грызунов. Продолжающееся снижение численности малых сусликов, увеличение численности малых песчанок, мышевидных грызунов и их блох изменило эпизоотологическую обстановку по чуме в этом регионе (Тихомиров, 1991; Козлова, 1994; Попов с соавт., 1995; Яковлев, 1996). На юге Калмыкии и правобережья Астраханской области на территории Прикаспийского песчаного очага чумы в 2001 году на фоне широкой экспансии общественной полевки было выделено 5 штаммов возбудителя чумы от этих грызунов и собранных с них блох (Матросов, Кузнецов, Слудский с соавт., 2003). Периодические вспышки численности
107
и расширение их ареала отмечались здесь и ранее (Орлов, 1928; Огнев, 1950; Павленко, 1959).
В мае-июне 2001 г. нами были проведены специальные полевые исследования в северной части очага: было отработано 1174 ловушко- ночей, пройдено 15 маршрутов общей протяженностью 91 км, раскопано 35 «колоний», добыто 31 гнездо, отловлено 302 экземпляра общественной полевки и собрано 364 блохи. Особое внимание уделялось изучению интенсивности норовых и паразитарных контактов в многовидовых сообществах мелких млекопитающих. В этих целях помимо маршрутных учетов осуществлялся тотальный вылов зверьков на колониях полевок в 14 пунктах.
Известно, что для общественных полевок характерен «пульсирующий» тип многолетней динамики пространственной структуры популяций (Флинт, 1977). Несмотря на выраженную оседлость зверьков и приуроченность устойчивых поселений к локальным участкам стаций переживания, в благоприятные годы могут происходить вспышки размножения и активное расселение молодых зверьков за пределы резерватов (Золотухина, 1980; Касаткин, 1997). Формирование полноценных устойчивых во времени поселений этих полевок на новых территориях происходит, как правило, в течение нескольких сезонов и лет только после образования устойчивой пространственно-этологической структуры их группировок (Щипанов, Касаткин, 1996; Шилова, Касаткин, 2000). На вновь заселяемой территории общая численность зверьков может возрастать в десятки раз. Общественная полевка при этом может занимать доминирующее положение в большинстве местообитаний (Шилова с соавт., 2000).
Уровень численности популяций млекопитающих зависит от интенсивности размножения и темпов смертности в неблагоприятные периоды года. Полиэстральные виды грызунов способны размножаться круглый год, но в аридных условиях приносят потомство только весной и
108 осенью (Козлов, 1965). Общественные полевки во влажные годы дают 4-5 пометов в год, в засушливые - лишь 1-2. Лимитирующим фактором при размножении является наличие витаминных зеленых кормов (Хашаева, 1993). Интенсивному размножению способствует влажное лето предыдущего года с обилием сочной растительности и бурная вторичная вегетация осенью (Касаткин, 1997). Весеннее размножение начинается в марте и растянуто до мая. Осенью в зависимости от погоды и кормовых условий беременные самки начинают встречаться в конце августа - сентябре (Касаткин и др., 1998). У общественных полевок репродуктивная функция реализуется только в устойчивых, сформировавшихся группировках (Щипанов, Касаткин, 1996). Внутрипопуляционные механизмы регуляции численности у этого вида связаны в первую очередь с подавлением генеративной активности и снижением жизнестойкости зверьков (Емельянов, Михалевич, 1975).
Устойчивые и относительно крупные по площади поселения общественной полевки к 2001 г. сформировались в Приморском и Черноземельском ландшафтных районах. В Ильменном районе нами регистрировались лишь единичные колонии или группы колоний несложного строения. Такое распределение по заселяемой территории объясняется природными условиями. На Черных землях преобладают сплошные массивы целинных или залежных земель с рыхлыми почвами мелкобугристых или равнинных песков и супесей. Здесь полевки имеют возможность строить долговременные сложные норы-колонии и активно расширять ареал поселений. Последние распределяются в виде крупноостровных скоплений, насчитывающих от нескольких десятков до сотен колоний. Площадь таких поселений достигала нескольких сотен га. Для Приморского района характерны мелкоостровные и ленточные поселения с несколькими десятками колоний на площади до 100 га. В Ильменном районе с выраженной мозаикой ландшафтных фаций
109 общественная полевка создавала небольшие по площади островные и узкие ленточные поселения с малым количеством колоний (до 10).
Анализ архивных данных показал, что численность общественной полевки в Прикаспийском песчаном очаге за последние 50 лет никогда не поднималась до такого высокого уровня. В таблице 4 приводятся материалы за 1999-2002 гг., которые свидетельствуют о том, что к весне 2001 года численность этого вида в природных биотопах по сравнению с многолетней нормой увеличилась в 10 раз. Следует иметь в виду, что при стандартных учетах на ловушко-линиях улавливается лишь общая тенденция динамики численности этих зверьков (полевки плохо берут хлебную приманку). Материалы, приведенные в таблице 4, были дополнены данными маршрутного учета, более объективно отражающего уровень численности и долю этих полевок в структуре сообществ грызунов. Общественные полевки отмечались на всех 15 пунктах учета, на 10 из которых они явно преобладали по численности. В Черноземельском районе индекс доминирования оказался максимальным — 90%, в Приморском и Ильменном районах — 71 и 53% соответственно. Общая заселенность территории этими грызунами составила 5.5% (от 3% до 11 %).
Таблица 4. Динамика численности общественной полевки в открытых биотопах в Прикаспийском песчаном очаге чумы в 1999-2002 гг.
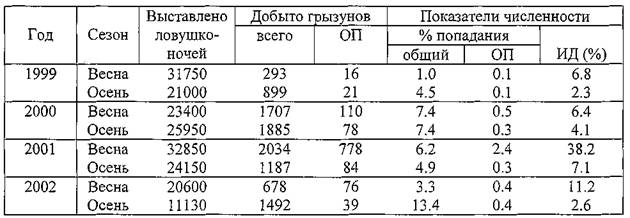
Примечание. ОП — общественная полевка; ИД — индекс доминирования общественных полевок.
по
На маршрутах получены интересные материалы о стадиальном размещении грызунов (таблица 5). Общественная полевка встречалась во всех типах ландшафтных фаций. Колонии обнаруживали даже на заливных лугах и солончаковых низинах с галофитами. Охотнее всего зверьки селились в зарослях кустарников: тамарикса, селитрянки, джузгуна и на сухом разнотравье, где при заселенности 12.8% плотность полевок составила 27.4 особей на га. Также охотно они занимали антропогенные биотопы: валы, насыпи, склоны каналов, обочины дорог, поросшие бурьяном. Здесь заселенность составила 9.4%, а плотность — 19.0 особей на га. Оптимальными для обитания оказались сухие разнотравные целинные и залежные степи, занимающие большие пространства. В мае- июне 2001 г. в пунктах учета наряду с обитаемыми колониями полевок встретились и 33 нежилые, что составило 6.2 % от общего числа регистрируемых. Отмечались и строящиеся колонии, которые на новых территориях всегда обнаруживались в оптимальных биотопах (кустарники, валы, насыпи). Общественные полевки стали проникать в жилые дома и скирды.
Обобщая полученные данные, можно констатировать, что емкость оптимальных для обитания общественных полевок биотопов позволяла зверькам и далее расселяться на очаговой по чуме территории. Вместе с тем известно, что в популяциях этого вида большое значение имеют процессы авторегуляции численности в местах постоянного обитания и малозначимы процессы миграции. Для выживания вида более важно поддержание стабильной численности полевок на небольших участках резерватов (Щипанов, Касаткин, 1996).
Общественные полевки проводят большую часть жизни в норе (Огнев, 1950). Они строят сложные норы-колонии, открывающиеся на поверхности земли множеством норовых отверстий (Воронов, 1935). В отличие от обыкновенной, общественная полевка селится группами в ксерофильных биотопах, избегая участков с избыточным увлажненем.
Таблица 5. Численность грызунов по типам ландшафтных фаций в северо-восточной части Прикаспийского песчаного очага чумы
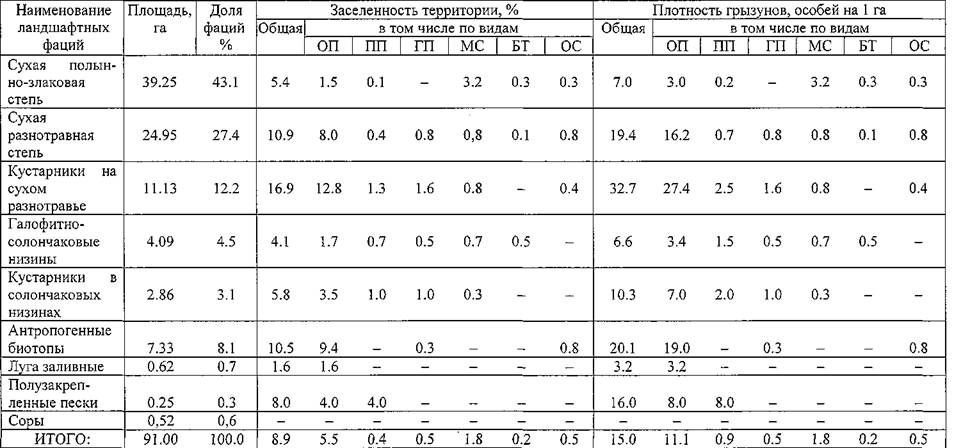
Примечание. ОП — общественная полевка, Illl — полуденная песчанка, ГП — гребенщиковая песчанка, MC — малый суслик, БТ — большой тушканчик, ОС — обыкновенная слепушонка.
112
Непригодны для строительства нор сыпучие песчаные почвы и крупнощебнистые грунты. Для этого зверьки выбирают участки с плотными сухими грунтами. При заселении пустующих территорий общественные полевки сначала устраивают простые норы с единичными выходами. Формирование устойчивых поселений со сложными норами- колониями достаточно длительный, многолетний процесс (Щипанов, Касаткин, 1996).
Массовое расселение общественных полевок на новых территориях в Северо-Западном Прикаспии происходило в течение 2-3 последовательных сезонов. Избегая сыпучих субстратов, зверьки все же предпочитали разрыхленные грунты антропогенного происхождения (залежи полей, насыпи, валы). Часто в качестве основы при строительстве колоний использовались подземные норы слепушонок, брошенные городки песчанок и сусликовины. Полноценные норы-колонии общественной полевки начали хорошо обнаруживаться на местности лишь осенью 2000 — весной 2001 гг. На открытой местности они идентифицировались по наличию большого числа входов (10-30), соединенных тропами, и по разреженному травостою. Краевые норы часто не связывались с центром и заканчивались слепо, соединяясь с 2-3 такими же, и служили для укрытия зверьков. Норовые отверстия располагались на расстоянии 0.5-3 м друг от друга на площади от 30 до 150 кв. м. Сами колонии в островных и ленточных поселениях размещались группами на расстоянии 10-50 м. В крупных кружевных поселениях соседствующие колонии сливались друг с другом и границы между ними оказывались неразличимыми. В кустарниках норы полевок отыскивались с большим трудом, число входов здесь сокращалось, а часть из них скрывалась от визуального наблюдения среди травы и корней.
В мае-июне 2001 года нами было раскопано 35 колоний общественной полевки. Каждая из них состояла из системы соединяющихся под землей ходов с несколькими кормовыми и 1-2
из
гнездовыми камерами. Наиболее посещаемые ходы, имеющие округлый профиль, приводили к гнезду. Кормовые камеры представляли собой расширенные, округлые в поперечнике полости по ходам нор. В период наших раскопок наполнение их никогда не было значительным (до 100 г) и состояло из зеленых остатков растений (стебли и колоски мортука, житняков, мятлика, рогоглавника, листья солянок, полыней, семена клоповника). В колонии обнаруживалась, как правило, одна гнездовая камера шарообразной формы диаметром 10-15 см. Она никогда не заканчивалась слепо и имела по 2-3 выхода наружу. Глубина залегания гнезд варьировала от 30 до 50 см. Длина ходов, ведущих к гнезду, составляла не менее 1.5 м (максимально до 4 м) от наружного отверстия. На дне гнездовой камеры располагалось округлое плоское блюдцеобразное гнездо диаметром 8-10 см, выстланное измельченной травой. Строительным материалом часто служили злаки, реже полыни и прутняки. При раскопке колоний мы не обнаруживали гнезда с выводками и не встречали павших зверьков (более половины нор раскапывалось на эпизоотических по чуме участках).
В Северо-Западном Прикаспии специфическими паразитами полевок рода Microtusявляются блохи A. rossica, Nosopsyllus Consimilis, N. mokrzeckyi и Ctenophthalmus secundus(Иофф, Тифлов, 1954). По нашим данным в северной части Прикаспийского песчаного очага чумы на общественной полевке зарегистрировано 13 видов блох. Данные о видовом составе и численности эктопаразитов по материалам очеса зверьков в январе - мае 2001 г. приведены в таблице 6. Такие массовые сборы блох с этих зверьков получены здесь впервые. Индекс обилия в шерсти полевок оказался максимальным в феврале - марте и составил в среднем 1.5. При явном доминировании специфических блох мышевидных грызунов (94 %) обращает на себя внимание присутствие песчаночьих блох N. Iaeviceps, X. Conformisи сусликовых блох N. setosaи С. tesquorum,являющихся эффективными переносчиками чумы. Особенно важно отметить находки
114 на полевках блох P. irritans.К маю наблюдалось снижение численности блох самого массового вида A. rossica,связанное с фенологией их годового цикла.
Таблица 6. Видовой состав и численность блох, собранных с общественных полевок в Прикаспийском песчаном очаге чумы в 2001 г.
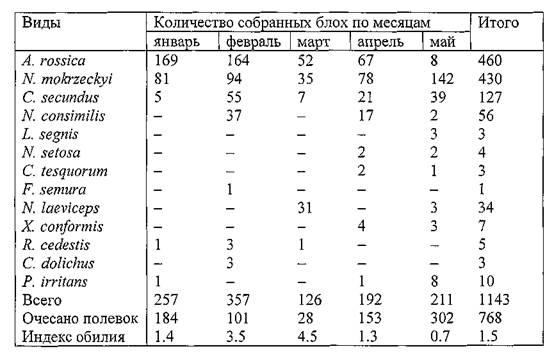
По материалам раскопок 31 гнезда общественных полевок в мае - июне 2001 г. было собрано 153 блохи: 65 — N. mokrzeckyi, 68 — Ct. secundus, 10 —X. Conformis, 6 —A. rossicaи 4 —N. Iaeviceps.Единичные блохи обнаруживались по ходам нор и в кормовых камерах (1-6), а большинство норовых блох было собрано в гнездах. Индекс обилия блох в гнездах составил 5.5. В гнездовой выстилке при разборе субстрата в этот период отмечалось большое количество личинок блох.
По материалам маршрутных учетов, облова колоний общественной полевки и паразитологических сборов отмечаются тесные норовые и паразитарные контакты этого вида с другими мелкими млекопитающими. На маршрутных учетах повсеместно регистрировали смешанные поселения полевок с гребенщиковой и полуденной песчанками, малым сусликом и обыкновенной слепушонкой (см. таблицу 5). На колониях
115 отлавливались домовая мышь, серый хомячок, обыкновенная полевка, полуденная и гребенщиковая песчанки, часто регистрировались следы посещений степными хорями и корсаками, численность которых в настоящее время высока в связи с многолетним запретом охотничьего промысла. В шерсти и гнездах общественной полевки обычны специфические блохи песчанок, сусликов и хищных млекопитающих (см. таблицу 6). По данным о видовом составе и численности блох на других грызунах отмечается, что массовые виды специфических блох полевок часто встречаются на основных носителях чумы — песчанках и сусликах. Такой широкий обмен эктопаразитами, по-видимому, здесь — явление обычное.
Эпизоотологическое значение общественной полевки в равнинных очагах чумы невелико (Эйгелис, 1973; 1980). Впервые возбудитель чумы выделен от этих зверьков в 1914 г. в казахстанском Зауралье (Кольцов, 1926). Единичные находки зараженных общественных полевок и счесанных с них инфицированных чумой блох обнаруживались в очагах Кавказа и Закавказья (Гниловская, 1991). В Северо-Западном Прикаспии в 1947 и 1948 гг. было выделено по 1 культуре чумы от блох N. setosa, добытых в гнездах общественных полевок (Павленко, 1959). Отмечается высокая устойчивость этих полевок к чуме, что подтверждено экспериментальными работами (Мамедзаде, 1948; Сардар, 1956).
Северо-восточная часть Прикаспийского песчаного очага чумы с 1979 г. по настоящее время остается одним из наиболее активных участков стойкого проявления чумы. Несмотря на изменение соотношения и численности фоновых видов грызунов-носителей, эта тенденция сохраняется. Снижение численности малых сусликов, вызванное естественными процессами циклического характера и антропогенной трансформацией природных ландшафтов, привело к снижению их роли в развитии эпизоотий. Основными носителями чумы выступили малые песчанки, численность которых в условиях опустынивания зональных
116 ландшафтов увеличилась и стабилизировалась (Тихомиров, 1991; Козлова, 1994; Яковлев, 1996).
В результате эпизоотологического обследования в первой половине 2001 г. в Прикаспийском песчаном очаге чумы было выделено 27 штаммов возбудителя чумы. Несмотря на низкую численность сусликов, большая часть штаммов изолирована от этих зверьков (11) и собранных с них блох (3). От блох, счесанных с малых песчанок, выделено 8 культур чумы. На фоне резкого подъема численности общественной полевки в их поселениях выделено 5 штаммов возбудителя чумы: 3 — от специфических блох полевок A. rossicaи N. mokrzeckyi, 1 — от песчаночьей блохи N. Iaevicepsи 1 — от самого зверька (через биопробу). Несмотря на повсеместное распространение и высокую численность общественной полевки, массового вовлечения ее в эпизоотический чумной процесс не произошло. Это хорошо укладывается в сложившиеся представления о малом эпизоотологическом значении этих полевок и их блох в равнинных очагах чумы: сами зверьки резистентны, а их специфические блохи плохо блокируются и передают чуму. Находки инфицированных полевок и их блох объясняются интенсивными норовыми и паразитарными контактами с основными носителями чумы — сусликами и песчанками. Последующее сокращение численности общественной полевки, вызванное авторегуляторными механизмами и неблагоприятными погодными условиями, привело к восстановлению прежнего, невысокого эпизоотологического статуса этого вида грызунов.
Причины высокой эпизоотической активности Прикаспийского песчаного очага чумы полностью не расшифрованы. Наблюдаемые сукцессии в зональных ландшафтах, связанные с естественными циклическими процессами и антропогенным воздействием, изменяют биоценотическую структуру природного очага чумы. Глубокая депрессия состояния популяции малого суслика, некоторая стабилизация численности малых песчанок сопровождаются расширением ареала
117 мышевидных грызунов. Многократное увеличение численности и широкое распространение общественной полевки в 2000-2001 гг. привело к тому, что этот зверек на непродолжительное время стал фоновым, а на некоторых территориях доминирующим по численности. В такие периоды общественная полевка может играть определенную роль в развитии чумного эпизоотического процесса. Однако уже к осени 2001 г. произошло повсеместное снижение численности общественной полевки до среднего многолетнего уровня. На основании проведенных исследований можно констатировать, что в настоящее время нет аргументированных оснований к изменению тактики эпизоотологического обследования на чуму Прикаспийского песчаного очага, обусловленному колебаниями его биоценотической структуры.
Здесь, как и в других очагах, большое эпидемиологическое и эпизоотологическое значение имеют синантропные грызуны как таковые и особенно их экзоантропные популяции. Домовая мышь, достигающая по валам оросительных каналов в зоне поливного земледелия 40-60% попадания на ловушко-линиях, в отдельные годы обеспечивала получение большого числа культур чумного микроба. Тесно контактируя с обитающими по соседству песчанками, домовая мышь вовлекается в эпизоотический процесс и может служить источником заражения чумой человека. Экзоантропные поселения этого грызуна в пределах эпизоотических участков должны подвергаться первоочередной дератизации. Наличие в очаге поселений серой крысы не только в жилье человека, но и в природных биотопах, также имеет большое эпидемиологическое значение. Более того, одна культура возбудителя чумы была получена от серой крысы, добытой на полевом стане в 1988 г. Ранее зараженных крыс отлавливали в 1936, 1938 и 1946 гг.
На грызунах, обитающих в очаге, паразитируют 27 видов блох, основными переносчиками выступают блохи N. Iaeviceps, N setosaи С.
118
tesquorum.Немаловажную роль в распространении инфекции играют блохи домовых мышей N. mokrzeckyi.
В прошлом (до 1954 г.) большинство культур чумы выделялось от малых сусликов и их специфических блох N setosaи С. tesquorum. Начиная с 1979 г., когда число сусликов в отловах резко уменьшилось, возбудитель обнаруживался при исследовании песчанок, мышевидных грызунов и их блох. В эпизоотический процесс вовлекались многие виды блох: N. Iaeviceps, N. setosa, С. tesquorum, С. pollex, F. semura, М. hebes, X Conformis, N. mokrzeckyi, N. consimilis, A. rossica.
Численность блох в Прикаспийском песчаном очаге чумы характеризуется в целом низкими показателями. Малая численность высоко активного переносчика — блохи X. conformis,либо ее полное отсутствие объясняют довольно вялое течение эпизоотий среди песчанок. Этому же способствует наличие значительных разрывов в поселениях прокормителей. Индексы обилия самой массовой блохи песчанок N. Iaevicepsна полуденных песчанках держатся на уровне 0.1-2.0, опускаясь в летние месяцы практически до 0. На гребенщиковых песчанках индексы обилия выше (2.0-4.0) и порой достигают 10.0-12.0 экз. блох на одном зверьке. В гнездах песчанок обнаруживается небольшое количество блох, а их миграция к устьям нор также выражена слабо (Васильев, Гниловская, 1989).
Наиболее значимой в эпизоотологическом и эпидемиологическом смысле является территория, примыкающая к дельте Волги (рукав Бахтемир) и берегу Каспия между дельтой Волги и устьем р. Кумы. Эта территория включает часть Кумского района и Приволжских песков, Приморский, Ильменно-придельтовый и Нарын-Худукский районы, а также восточную часть Черных земель. Ежегодная регистрация здесь зараженных животных позволила провести детальную оценку устойчивости эпизоотических проявлений на отдельных участках и на этой
119 основе осуществить эпизоотологическую дифференциацию наиболее активной части очага (Кузнецов с соавт., 2001).
Наиболее устойчивые эпизоотические проявления, начиная с 1981 г., были приурочены к зоне орошаемого земледелия, а точнее к стыку целинных земель и сельскохозяйственных угодий, оборудованных оросительными каналами и водогонами, берега которых часто зарастают бурьяном. Характер биоценотической структуры участков очаговости, подвергшихся антропогенной трансформации ландшафтов, также претерпел определенные изменения. Обитающие там мышевидные грызуны, численность которых и раньше достигала порой высоких значений, часто вовлекаются в эпизоотический процесс. Их поселения граничат с поселениями песчанок, а кое-где и сусликов. Отмечаемые многими авторами преобразования природы очага могли быть причиной его функционирования в постоянном режиме. Существенным является также то, что в годы с высокой эпизоотической активностью или в непосредственно следующие за ними, зараженные животные в очаге встречались не только в зоне орошаемых земель, но и далеко за их пределами (Васильев, Дятлов, 1987; Денисенко с соавт., 1987), а в годы с низкой активностью — практически только в их границах.
120
3.3.3.
Еще по теме Естественная и антропогенная трансформация Прикаспийского песчаного очага чумы.:
- Структура Прикаспийского песчаного очага чумы
- Структура Волго-Уральского песчаного очага чумы.
- Прикаспийский песчаный очаг (43).
- Ландшафтная приуроченность элементов пространственной структуры популяций песчанок в ВолгоУральском песчаном очаге чумы.
- Волго-Уральский песчаный очаг(16)
- Трансформация
- Лекция 3. «Медико-социальная характеристика природных и антропогенных катастроф»
- Трансформации энергии.
- Трансформация
- Особенности формирования и ранней диагностики аутоиммунного тиреоидита у детей в условиях повышенной антропогенной нагрузки
- 2.2.1 Формирование первичного очага
- 9.3. Трансформация клеток Escherichia coli.
- Реакция бластной трансформации лимфоцитов (РБТЛ)
- 12.2.2. Характеристика очага ядерного поражения
- б.Трансформация продуктов разрыва кольца в полупродукты цикла Кребса.
- ГЛАВА 39 РАК из невыявленного первичного очага
- Лечение первичного очага.