Ландшафтная приуроченность элементов пространственной структуры популяций песчанок в ВолгоУральском песчаном очаге чумы.
Под пространственной структурой популяций понимают закономерное размещение особей и их группировок по отношению к определенным элементам ландшафта и друг к другу (Шилов, 1977). Закономерное размещение особей определяет наиболее эффективное использование ресурсов среды — кормовых, защитных, микроклиматических и др.
Другое значение пространственной структурированности популяций заключается в . поддержании необходимого уровня внутрипопуляционных контактов между особями, которые обеспечивают видовые и биоценотические функции популяции (размножение, расселение, участие в круговороте веществ, энергии и др.). Грызунам — обитателям открытых пространств характерен преимущественно оседлый образ жизни, определяющий интенсивный тип использования пространства, при котором отдельные особи или их группировки в течение длительного времени эксплуатируют ресурсы ограниченной территории (Шилов, 1977). По сути, пространственная структура популяции представляет собой систему индивидуальных участков обитания животных. Некоторое количество участков обитания (Миронов, 1980), агрегированных до такой концентрации, при которой обеспечивается устойчивое общение особей, называется поселением. Предлагалось также такую группировку в совокупности с индивидуальными жилищами (норами), обладающую общностью всего комплекса биологических параметров, называть мерусом или элементарным поселением (Флинт, 1977). Важной характеристикой любого поселения служит взаиморасположение составляющих его нор, колоний, которое определяет его тип. По мнению Н.П.Наумова (Наумов,
74
1954), существует два типа поселений: диффузные и мозаичные. Главной причиной, определяющей формирование того или иного типа поселений на конкретной территории, является ее ландшафтная структура. Иными словами пространственная структура животного населения, так же как и эпизоотологическая структура очага чумы, в основном соответствует ландшафтным, геоботаническим или иным природным подразделениям территории (Гайский, 1930; Флинт, 1977; Коренберг, 1979).
Целенаправленное и обоснованное изучение пространственной структуры популяций носителей и переносчиков в аспекте их разнокачественности осуществлено рядом авторов в природных очагах Сибири (Зонов с соавт., 1988; Вержуцкий, 1990; Вержуцкий с соавт., 1994; Чипанин, Попков, 1997).Поселения определенного вида животных (не зависимо от их размеров и количества), расположенные в пределах единой территории, внутри которой отсутствуют рассекающие ее протяженные экологические барьеры, обычно объединены также в единое сообщество, называемое популяцией. Внешняя граница такой территории, служащая границей ареала конкретной популяции, обязательно должна обладать свойством, физически затруднять контакты между животными, обитающими по обе ее стороны (Свирежев, 1968). При этом речь не идет о полном прекращении контактов, а лишь об их относительном ограничении.
Методом изучения распространения и размещения животного населения является зоологическое картографирование. Любые исследования пространственной структуры популяций, биохорологической структуры вида и других особенностей использования территории животными невозможны без применения картографического метода (Исаков, 1963; Кучерук, 1965; Кучерук с соавт., 1965; Тупикова, Комарова, 1979; Бокштейн, Дубровский, 1981).
Принципы построения карт группируются в два основных направления (Коренберг, 1979). Первое из них предполагает
75 картографирование на геоботанической или ландшафтной основе, при котором зоологические данные, полученные в пределах определенных выделов, могут быть экстраполированы на всю типологически сходную территорию (Попов В.П., 1978; Матросов с соавт., 1984; Эйгелис с соавт., 1989). Этот принцип построения карт называют типологическим. Второе направление основывается на представлении, что животное население имеет собственную структуру, которую выявляют отдельно (независимо) от разнородности территории. Такой путь построения карт назван дифференциальным.
Один из наиболее важных и сложных вопросов зоологического картографирования связан с экстраполяцией данных, получаемых из ограниченного числа мест, на участки, не подвергшиеся прямому обследованию (Коренберг, 1979).
К сожалению, человечество еще не в состоянии предложить метод сплошного картографирования животных на больших территориях, позволяющий вообще обойтись без какой-либо экстраполяции, что вынуждает исследователей разрабатывать ее принципы и практические приемы.Основа оптимальной экстраполяции данных была заложена нами еще при выборе метода картографирования, который мог обеспечить получение наиболее репрезентативных и рандомизированных материалов. Первое условие (репрезентативность) могло быть выполнено при использовании маршрутного метода картографирования (Ротшильд с соавт., 1975), второе (рандомизированность) — при предварительном вычерчивании на рабочей картографической основе параллельных прямолинейных маршрутов, располагаемых строго через равные метрические промежутки, что полностью исключает влияние субъективного фактора. Этим целям в полной мере отвечал маршрутный метод, использованный при картографировании поселений песчанок Восточного Закавказья (Эйгелис с соавт., 1979).
76
По нашему мнению, в основе крупномасштабного зоологического картографирования должен лежать принцип, совмещающий в себе и типологическое, и дифференциальное направления. Приемом детализации графической информации был избран формально-территориальный способ ее отображения, сочетающийся с внутриконтурной экстраполяцией. Применение этих приемов позволило нам получить достаточно убедительные результаты обработки данных полевого картографирования.
Необходимым условием успешного картографирования является выбор и конкретизация объекта съемки. В Волго-Уральских песках этот вопрос уже рассматривался исследователями (Ротшильд с соавт., 1975), однако отсутствие единого мнения о количественной оценке и вообще о характеристике городка песчанок как объекте картографирования, потребовало рассмотреть его вновь и сделать необходимые уточнения.
Наименьшей (элементарной) пространственной единицей, по которой регистрируется наличие полуденных и гребенщиковых песчанок на той или иной территории и определяется их численность, является городок.
Городком называют систему подземных ходов («нору») этих песчанок, с одним, как правило, используемым гнездом и с несколькими (до трех десятков) выходами на поверхности земли (Ротшильд с соавт., 1975; Бурделов с соавт., 1981; Скиртачев, 1981; 1985; Постников, Ротшильд, 1985). Обычно все ходы «молодой» норы сообщаются между собой под землей, однако, в «старых» городках могут быть и изолированные части, именуемые защитными, временными, кормовыми и т. д. норами (Ралль, 1931; 1938; 1941). Городок, оконтуренный на поверхности земли по крайним норовым отверстиям, вписывается в квадрат 10?10 м и является жилищем одной (как правило) взрослой песчанки (Скиртачев, 1985). Из этого описания следует, что городок — это наименьший неделимый элемент пространственной структуры ареала (популяции) малых песчанок Волго-Уральских песков. Именно городок,
77
как поверхностный эквивалент подземной норы, является главным объектом полевой съемки.
В ряде случаев, особенно при высокой численности песчанок, городки отдельных зверьков могут располагаться в непосредственной близости друг с другом. По взаиморасположению норовых отверстий на поверхности земли бывает трудно определить, одному или нескольким городкам они принадлежат. В реальных условиях песчаной пустыни идентификацию отдельных городков осуществляют, применяя разработанное нами правило (Кузнецов с соавт., 1982). Если группа норовых отверстий занимает площадь более 100 м2 или между крайними отверстиями вдоль наибольшего из поперечников данной группы расстояние превышает 15 м, то считают, что в этом месте имеется больше одного городка. Во всех подобных случаях каждые 100 м2 группы относительно равномерно расположенных отверстий принимают за один городок, а остаток, превышающий эту площадь на 20 м2 и более — еще за один. Небольшие отклонения (до 20 м2) могут быть округлены. Это правило корректируют в случаях заметной неравномерности расположения входов нор на поверхности земли, особенно со сложным рельефом, указывающей на физические разрывы в системе подземных ходов.
В природе городки располагаются на любом расстоянии друг от друга, начиная от непосредственного «соприкосновения» и дальше. Группа городков, в которой расстояние между соседними в любой паре не превышает предельную дистанцию, при которой еще сохраняется возможность регулярных контактов их обитателей друг с другом, называется поселением, население которого можно отождествить с парцеллярной группировкой (Чекашов с соавт., 2001). Сложность заключается в установлении размера такой дистанции. По нашим данным (см. Главу 4), среднее расстояние ежесуточных перемещений полуденных и гребенщиковых песчанок в Волго-Уральских песках составляет приблизительно 70 м (85 м весной и 50 м осенью), наибольшее — 220-260
78
м. Учитывая эти данные, мы определили, какие средние дистанции между городками являются пороговыми и какие уровни агрегированности нор они разграничивают.
Удовлетворительного ответа на вопрос о расстояниях между норами, исключающих прямые контакты их обитателей (песчанок), в доступной литературе мы не обнаружили, поэтому рассмотрим его с использованием простых логических построений. Известно, что норы (колонии, городки и т. д.) грызунов располагаются на территории неравномерно, причем степень неравномерности может весьма сильно различаться. Дислокация каждого конкретного городка обусловлена многими обстоятельствами, среди которых элементарная случайность играет не последнюю роль. При увеличении численности животных расстояния между их норами сокращаются, и вариабельность этих расстояний уменьшается.
Анализ модельных ситуаций логично проводить, если задано условие, при котором все расстояния между соседними городками одинаковы. Это допущение предполагает размещать норы в узлах правильной треугольной решетки, образованной сомкнутыми равносторонними треугольниками, что обеспечивает наиболее плотную «упаковку» точек (вершин треугольников) на плоскости (Кудрин, Завалишин, 1973; Эфрос, 1982). Естественно, что рассматривать ареал вида или отдельной популяции как одну сплошную решетку нереально.
А вот одно поселение грызунов (как небольшая часть популяции) вполне может быть представлено цельным фрагментом треугольной решетки. Параметры такого модельного поселения могут соответствовать средним характеристикам естественного и употребляться для расчета и оценки недостающих величин. Поясним это на следующем примере.Для учета численности (плотности) малых песчанок используют метод подсчета площади, занимаемой их городками, и доли этой площади (в %) на учетном трансекте по правилам, описанным в разделе 2.2. (Кузнецов с соавт., 1982;). При равномерном размещении городков
79
обитателю каждого из них будет принадлежать «индивидуальный участок» в виде правильного шестиугольника (рис. 4), поскольку треугольная и шестиугольная решетки дуальны друг к другу (Эфрос, 1982). Условность такого участка заключается в том, что если его обитатель передвигается лишь в его пределах, то со своими соседями (так же, как и с их норами) он не контактирует. Эта область, примыкающая к городку, скорее эквивалентна охраняемой «территории», однако, в случае с малыми песчанками таких территорий просто нет и поэтому шестиугольники следует рассматривать как «индивидуальную норму» земной поверхности, окружающей один городок и не имеющей других городков.
Доля заселенной площади внутри всего поселения будет соответствовать доле площади одного городка относительно площади «индивидуального участка» (шестиугольника). Зная «стандартизованную» площадь городка (So =100 м2) и долю (в %), которую он занимает на участке (заселенность — Z), можно вычислить площадь одного участка (S) и расстояние между центрами двух соседних участков (R)по следующим формулам: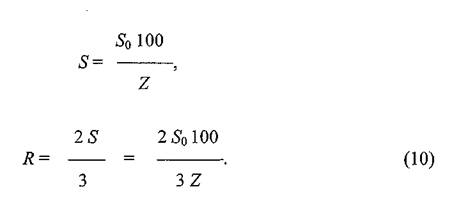
Среднее расстояние между городками в поселении служит важной его характеристикой, используя которую можно оценить оптимальность контактов между животными. Приняв среднее расстояние между жилищами зверьков в поселении, равным 70 м (см. раздел 4.1.), можно полагать, что практически каждая особь при ежесуточных перемещениях сможет достигать нор своих соседей.
80
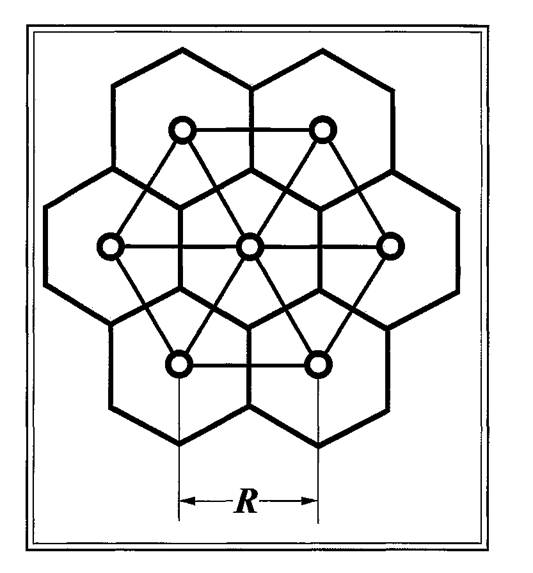
Рис. 4. Модельное поселение малых песчанок, состоящее из 7 городков со средним расстоянием между ними, равным R.
81
В поселении с расположением городков через 140 м число зверьков, свободно преодолевающих эту дистанцию, уменьшится. Тем не менее, норы мало- и среднеподвижных особей будут посещаться хотя бы одним (высокоподвижным) из шести его ближайших соседей. И в этом случае контакты среди обитателей поселения останутся достаточными. Если продолжить увеличение этого расстояния (за счет «разрежения» поселения), контакты будут плавно ослабевать, а предел их достаточности наступит, очевидно, при удалении городков друг от друга более чем на 220 м (максимальное суточное перемещение). Учитывая трудность точного и достоверного определения расстояния достаточных контактов и опасаясь невольного его завышения, считаем необходимым округлить в сторону уменьшения полученное в полевых экспериментах значение до 200 м.
Оптимальность контактов животных в поселении обеспечивается их стремлением располагать свои жилища (норы, колонии, городки) на самом удобном расстоянии друг от друга. Это стремление представлено двумя противоположными тенденциями — «притяжения» и «отталкивания», обеспечивающими как образование агрегаций животных, так и их расселение (Шилов, 1991 а; 1991 б). Стремление зверьков удаляться от своих соплеменников при устройстве нор на такое расстояние, при котором связь с ними не нарушена, а эксплуатация ресурсов наименее конкурентна, обеспечивает относительно равномерное (близкое к узлам правильной треугольной решетки) распределение городков. Это «центробежное» стремление действует до некоторого предела, после которого возрастает роль противоположного стремления — к сближению. В реальных условиях сближение не может в одинаковой мере и «геометрически правильно» реализовываться у всех членов поселения, поэтому равномерность расположения «сблизившихся» городков будет нарушена с учетом многих других причин подобного нарушения. Размеры поселения, разреженного до нижней грани оптимальной плотности, увеличиваться уже не будут (если не учитывать результаты размножения),
82 а равномерность дислокации нор в нем снизится. Реальная картина сближения может быть представлена группированием городков в более «густые» пятна, полосы, ленты и т.д. Крайний (абстрактный) случай — выстраивание нор в линию. В этом случае дистанции между городками в линии окажутся тем меньше, чем крупнее по площади было исходное поселение и чем больше городков было в нем, а длина линии станет адекватной поперечнику поселения. К сожалению, многие важные аспекты формирования пространственно-этологической структуры популяций грызунов изучены явно недостаточно (Шилов, 1991 а).
Естественно, что крайние варианты в природе не встречаются, а разные промежуточные стадии при непрекращающихся перегруппировках весьма разнообразны. Кроме того, на процессы сближения-расселения оказывают мощное модифицирующее воздействие многие природные факторы, такие, например, как рельеф местности, другие ландшафтные особенности конкретных участков, «кормность» угодий, их защитные свойства и т. д. Все это определяет фактическую неравномерность (мозаику) заселения территории грызунами, даже в так называемых диффузных поселениях (Наумов, 1954). По нашему мнению, всем грызунам открытых пространств свойственно внутреннее стремление образовывать равномерные диффузные поселения, а те или иные отклонения от такого типа использования пространства обусловлены преимущественно факторами «пятнистой» среды (Пианка, 1981).
При попытках смоделировать процесс перегруппировки нор песчанок «на бумаге» стало ясно, что наиболее легко и естественно возникает структура поселения, напоминающая сеть, когда одни сгущения городков связаны с другими посредством «цепочек» из таких же жилищ зверьков. Структура из разобщенных «пятен» могла возникнуть в случае резкого снижения количества городков и сохранения малого их числа на обширной территории. Однако здесь следует заметить, что подобные эксперименты на пространственных моделях могут быть осуществлены
83 качественно и на высоком уровне лишь профессиональными математиками, поэтому мы не делаем попыток углубленного анализа этого сложного вопроса.
Среднее расстояние между городками в поселении, вычисленное по заселенности территории, можно использовать для оценки оптимальности контактирования особей в нем при зарегистрированной в настоящий момент плотности населения. Если вычисленное расстояние превышает удвоенное предельное перемещение (2?200 м), высока вероятность, что на данном участке дислоцировано не одно цельное поселение, а несколько разобщенных. Реально существующие (или предполагаемые) разрывы между группами агрегированных городков, существенно превышающие 400 м, могут расцениваться как «разделительные пространства» между отдельными поселениями. Такие пространства в большинстве случаев весьма эфемерны и могут приниматься во внимание только лишь в период их обнаружения (не более одного-двух сезонов). Либо их разделительные свойства обусловлены особыми ландшафтными причинами, препятствующими заселению этих пространств песчанками. В любом случае их изолирующая роль не абсолютна, а дальнейший статус непредсказуем. В качестве примера можно привести «разделительную» функцию реки Урал, не мешающей регулярной экспансии больших и краснохвостых песчанок с левого берега на правый (Трофимов с соавт., 1974; Лавровский с соавт., 1978; Бурделов, Самарин, 1989; Самарин, Пономарев, 1989).
Приняв на вооружение способ вычисления среднего расстояния между городками и используя среднюю и максимальную дистанции ежесуточных перемещений песчанок от городка к городку, мы получили возможность предметно характеризовать плотность этих животных на любом участке местности, где получены данные об их численности. Для этого были разработаны пороговые значения плотности (заселенности) при
84
которых, по нашему мнению, должно меняться качество взаимоотношений между зверьками в поселении.
Среднее расстояние между городками, равное максимальному радиусу ежесуточных перемещений (200 м), соответствует достаточно низкой плотности песчанок и принято как одно из пороговых значений (фиг. 4 на рис. 5). При уменьшении этой дистанции сначала возникают спорадические, затем все учащающиеся норовые контакты между особями. При расстоянии, превышающем 200 м, контакты возможны лишь на поверхности земли («наземные» контакты, когда зверьки не достигают нор ближайших соседей). При разобщенности городков в 400 и более метров (наибольшая пороговая дистанция — фиг. 1 и 2 на рис. 5) прекращаются и эти контакты.
Напомним, что для анализа использованы абстрактные модельные ситуации, построенные на усредненных данных и представляющие собой «идеальный» случай. Выводы, сделанные на таких моделях, будут справедливы лишь для среднестатистической ситуации и могут значительно отличаться от отдельно взятых природных вариантов, учитывая их широкую вариабельность.
При средней разобщенности городков в 400 и более метров (фиг. 1 и 2 на рис. 5) считать их входящими в одно цельное поселение, по всей видимости, нельзя. Ассоциация городков, отстоящих друг от друга внутри нее более чем на 200 м, но не более чем на 400 (фиг. 3 на рис. 5), вероятно уже обладает признаками поселения. Практика убеждает в том, что пороговым значением, разграничивающим «поселение» и менее ассоциированную группировку, следует все же признать дистанцию между городками малых песчанок в 400 м. В пользу этой дистанции можно привести известные факты об увеличении показателей подвижности грызунов при снижении их численности
85
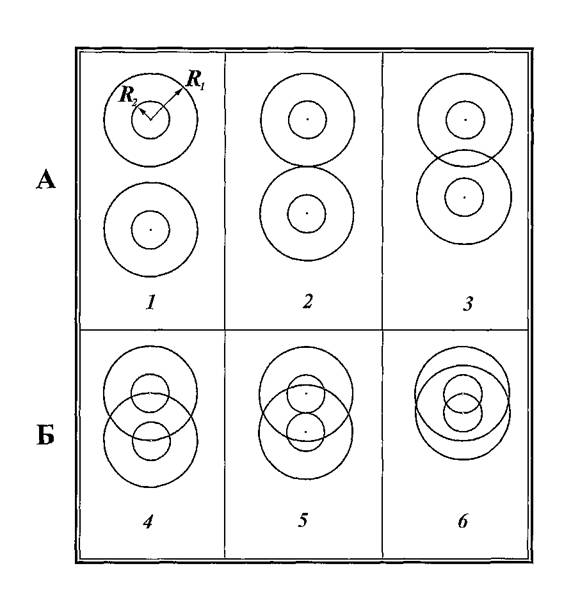
Рис. 5. Преобразование «наземных» (А) контактов зверьков в норовые (Б) при сокращении дистанции между городками. 1) R> 400 м; 2) R = 400 м; 3) 400 м >R > 200 м; 4) R = 200 м; 5) R = 140 м; 6) R = 70 м, где R — расстояние между городками. Радиус суточных перемещений: максимальный (7? і) = 200 м и средний (R2) = 70 м.
86
(плотности) и увеличении физической площади земли, приходящейся на одного зверька. То есть, чем дальше грызуны живут друг от друга, тем длиннее становятся их ежедневные перебежки или, иными словами, увеличивается радиус их участков обитания (Попов С.В. с соавт., 1989). Этому явлению можно дать несколько различных объяснений, которые, однако, не меняют его суть.
Диапазон плотностей, наблюдаемый при сближении городков от 200 до 140 м (удвоенное среднее расстояние ежесуточных перемещений), можно охарактеризовать как дистанции, обеспечивающие «слабые норовые контакты» (фиг. 4 и 5 на рис. 5). При плотностях, соответствующих такой взаимной удаленности городков, достигать нор соседей будут все подвижные особи. При дальнейшем уплотнении и уменьшении дистанций от 140 до 70 м (фиг. 6 на рис. 5) будут наблюдаться «субоптимальные норовые контакты», а затем (менее 70 м) — «оптимальные». Минимальная дистанция оптимальных контактов, при снижении которой начинается переуплотнение животных и возникновение избыточных («стрессирующих») контактов, нам не известна, поэтому чисто умозрительно и условно ее можно принять равной 35 м (половине среднего расстояния массовых перемещений).
Таким образом, мы установили четыре пороговые дистанции между городками в поселении, разграничивающие плотности грызунов на единицу площади, характеризующиеся определенным уровнем контактирования особей. Эти дистанции: 400, 200, 140 и 70 м. Пятая пороговая дистанция (35 м) может быть принята лишь условно и мы ее приводим как гипотетический прообраз такой дистанции. Зависимость заселенности территории поселений от пороговых расстояний между городками, приведена в таблице 3.
87
Таблица 3. Соотношение среднего расстояния между городками песчанок, заселенности территории и площади условного участка обитания
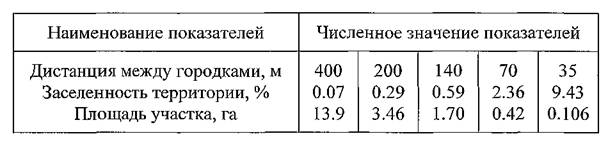
Заселенность территории весьма просто переводится в показатель численности, выражающийся числом зверьков на 1 га (см. разд. 4.2.). Для этого значение заселенности (в %) умножают на коэффициент плотности песчанок в городках. Весной этот коэффициент равен 0.7, осенью — 1.0. Используя данные таблицы 3 и вычисленные на ее основе приближенные показатели численности грызунов, получаем, что диапазон оптимальных и субоптимальных плотностей расположен в пределах от 0.5 до 7-8 зверьков на га, а возможно и несколько шире. По многолетним данным региональных противочумных станций и по публикациям известно, что численность полуденных и гребенщиковых песчанок в Волго-Уральских и Прикаспийских песках (по сумме двух видов) очень редко бывает ниже 2 особей на 1 га (Ралль, Демяшев, 1938; Павлов, Климченко, Лисицын с соавт., 1957; Павлов, 1961; 1962; 1965; Семенов с соавт., 1968; Павлов с соавт., 1977; Тлегенов с соавт., 1981). Из этого можно заключить, что практически всегда вся очаговая территория (не считая интразональных включений) имеет население грызунов, плотность которого соответствует оптимальному уровню внутрипопуляционного контактирования особей в любой его части. То же самое мы обнаружили на стационарных участках наблюдений, когда при достаточно низкой, казалось бы, численности песчанок (1 особь на 1 га), уровень норовых контактов и интенсивность обмена эктопаразитами были весьма высокими. Все это свидетельствует о том, что диапазон экологических условий, обеспечивающих возможность распространения возбудителя чумы по территории природного очага
88 достаточно широк. При наличии возбудителя в популяциях его носителей и переносчиков, эпизоотические проявления, скорее всего, были бы обнаружены. Если этого не происходит, то, очевидно, вообще отсутствует чумной микроб. Вероятно, в это время физиологические параметры носителей и переносчиков не обеспечивают размножение и передачу возбудителя, что приводит к его полной элиминации. Однако изложенное выше мнение может быть принято лишь при надежном и многократном его подтверждении.
Изучая пространственную структуру ареала полуденных и гребенщиковых песчанок в Волго-Уральском песчаном очаге чумы, мы попытались оценить реальность и продолжительность существования «ядер поселений» этих грызунов. Для этого в течение девяти сезонов (осень 1983 г., весна и осень 1984-1987 гг.) путем многократного картографирования изучали динамику элементов пространственной структуры ареалов песчанок на четырех стационарных участках, расположенных в различных частях изучаемого региона (Кузнецов с соавт., 1991). Один из участков располагался на западной кромке песков недалеко от урочища Тугайхудук, другой — на южной кромке песков в ур. Тойгенсор. Еще два участка (Истай и Бармак) находились в центральной части песков. Соседние участки отстояли друг от друга не менее чем на 40, крайние — на 110-150 км. Основными критериями выбора участков были их достаточная территориальная разобщенность, принадлежность к различным ландшафтным категориям (урочищам, местностям) и наличие различных по плотности и размерам поселений песчанок.
Ландшафтная структура юго-западной части песков, а также пространственная структура ареалов полуденных и гребенщиковых песчанок на этой территории были изучены нами в начале 80-х годов прошлого столетия (Матросов с соавт., 1984; 1987; Кузнецов с соавт., 1989; 1990; 1991; Эйгелис с соавт., 1989; Матросов, Кузнецов, 1994; 1996; Матросов, Кузнецов, Ерофеев, 1996; Матросов, Кузнецов, Скиртачев,
89 2001), что подробно отражено в диссертационной работе А. Н. Матросова (1992). Ландшафтно-геоботаническое дешифрирование аэрофотоснимков и проведенное на его основе зоологическое картографирование (Матросов с соавт., 1984; Эйгелис с соавт., 1989; Кузнецов с соавт., 1990; Матросов, Кузнецов, Ерофеев, 1996) позволили оконтурить участки сохранения поселений песчанок в депрессию численности на площади более 2 млн. га. Детальная оценка значимости ландшафтных фаций и урочищ в обеспечении устойчивости поселений зверьков стала возможной после проведения многократного картографирования четырех ключевых участков (Кузнецов с соавт., 1989; 1991).
В качестве картографической основы были использованы топографические карты масштабов от 1:25 000 до 1:100 000 и аэрофотоснимки масштабов 1:10 000 — 1:35 000. Основная форма использования аэрофотоснимков — фотосхемы, смонтированные на картоне в рамках трапеций, масштаба, близкого к масштабу залета (см. раздел 4.1.). Нанесение контуров ландшафтных элементов осуществляли на прозрачной лавсановой пленке с глянцевой (не матированной) поверхностью, которую накладывали на фотосхему. Процессу вычерчивания отдешифрированных контуров ландшафтных элементов предшествовал полевой этап выбора их эталона на небольшом числе фотосхем. В дальнейшем все дешифрирование проводили камерально по эталонам, подготовленным в поле (Вышивкин, 1977; Живичин, Соколов, 1980; Аковецкий, 1983). Перед дешифрированием ландшафтных элементов на фотосхемы наносили углы трапеций соответствующего масштаба (секторов). Затем в процессе фотокопирования каждого листа ландшафтной карты (сектора) использовали фототрансформацию проецируемого изображения и децентрацию негатива для устранения возникающих искажений (Лобанов, 1978; Мурашев, Гебгард, 1985;).
Для подсчета площадей ландшафтных элементов, имеющихся в каждом секторе, применяли полярный планиметр и прозрачную сеточную
90 палетку. Полученные указанным способом схемы распределения ландшафтных элементов и их количественные характеристики использовали в качестве основы при картографировании поселений малых песчанок, а также при эпизоотологическом обследовании территории. Изготовленные крупномасштабные ландшафтные карты позволили осуществить внутриконтурную экстраполяцию зоологической информации на ландшафтной основе (Матросов с соавт., 1984).
Картографирование поселений малых песчанок осуществляли маршрутным методом (Эйгелис с соавт., 1979), соблюдая общепринятые для таких работ правила (Господинов, Сорокина, 1974; Ганьшин с соавт., 1977; Руководство по топографической съемке ..., 1977; Грюнберг с соавт., 1991). Для этого на территории участка разбивали створы параллельных маршрутов, отстоящих друг от друга на 2 км, длиной 10-20 км. Суммарная длина маршрутов на всех участках равнялась приблизительно 480 км. Единицей отсчета длины (а также площади) трансекта служил его стометровый отрезок (0.25 га при ширине 25 м), отмеряемый по спидометру автомашины. Учетчик подсчитывал число встреченных городков песчанок на каждой стометровке, независимо от того, располагались ли они вразброс или в непосредственном соприкосновении друг с другом. За городок, как уже говорилось, принимали скопление норовых отверстий, вмещающееся в площадку 10?10 м. Явно нежилые городки не учитывали.
При обработке полученных данных картографические маршруты разбивали на километровые отрезки. Поскольку все они жестко фиксированы на местности, то имелась реальная возможность следить за численностью песчанок на каждом отрезке. Данные о заселенности (доля площади трансекта, занятая городками, выраженная в процентах) каждого километрового отрезка экстраполировали на прямоугольную ячейку площадью 2 км2 — по одному с каждой стороны от оси маршрута. В каждую полученную таким образом формально-территориальную ячейку
91
вписывали значение заселенности ее конкретным видом песчанок (полуденная или гребенщиковая). В результате на каждый участок и вид грызуна было изготовлено по девять хронологических схем дислокации формализованных поселений песчанок, характеризующихся той или иной плотностью зверьков.
Применительно к результатам картографирования и в соответствии с нормативами ассоциированности нор (см. таблицу 3) поселением песчанок следует считать группу их городков, среднее расстояние между которыми не превышает, по меньшей мере, 200 м. Заселенность поселения в соответствии с этим расстоянием составляет 0.29% его площади. Территорию, в меньшей степени заселенную песчанками, очевидно, нельзя считать занятой одним цельным поселением. При анализе формальнотерриториальных карт за поселение принимали одну или группу соприкасающихся стандартных ячеек, заселенность каждой из которых была не менее 0.29%.
В целом период картографирования пришелся на продолжительный период депрессии численности малых песчанок. Средняя заселенность полуденными песчанками любого из трех участков — Бармак, Тойгенсор и Тугайхудук — ни разу не превысила 3.8%, гребенщиковыми — 2.4%. На участке Истай заселенность была выше: до 9.8% — полуденными и 7.7% — гребенщиковыми песчанками. Наиболее низкими показателями численность песчанок характеризовалась в весенние сезоны 1984, 1985 и 1987 гг. Анализ именно этих и смежных с ними сезонов позволил получить представление о сохранении поселений песчанок в периоды ярко выраженной депрессии численности, об их размерах, плотности зверьков в них и приуроченности к одному и тому же месту.
На участке Бармак заселенность полуденными песчанками была минимальной в весенние сезоны 1985 и 1987 гг. (0.15%). Заселенность отдельных формально-территориальных ячеек не превышала 1.6%. Из 162 ячеек весной 1985 г. были заселены 38, весной 1987 г. — 45. В первом
92
случае заселенные ячейки располагались единично или группами (по 2-7). Во втором случае большинство ячеек были собраны в две группы из 12 и 22 ячеек. Из 38 заселенных весной 1985 г. ячеек вторично заселенными . весной 1987 г. оказались только 14, а остальные (31) располагались, как правило, рядом с ячейками, заселенными в прошлую депрессию численности. Об отсутствии строгой территориальной приуроченности сохраняющихся в разные периоды депрессии поселений полуденных песчанок говорит и тот факт, что из 162 ячеек на участке только две имели городки этих грызунов в течение всех девяти сезонов картографирования. Численность гребенщиковых песчанок на участке Бармак была постоянно низкой. Плотных поселений этого грызуна не обнаружено там не только в депрессивные, но и в более благоприятные сезоны.
На участке Тугайхудук отмечены те же закономерности в динамике поселений полуденной песчанки, что и на участке Бармак. Так, в период депрессии их численности весной 1984 г. там оставались заселенными 16 ячеек из 72. В следующий период депрессии весной 1987 г. таких ячеек было 22, причем повторно заселенных — только 4. Весьма сходная картина динамики поселений полуденных песчанок наблюдалась и на участке Тойгенсор. Достаточно крупных поселений гребенщиковой песчанки не обнаружено ни на том, ни на другом участке. На них не отмечено ни одной ячейки, постоянно заселенной одним из видов песчанок.
Учитывая крайнюю неустойчивость поселений песчанок вблизи кромки песков, как на юге, так и на западе массива (участки Тойгенсор и Тугайхудук), было решено более детальный анализ провести только по участкам в центральной части Волго-Уральских песков. Считая установленной для эпизоотологии чумы в этом регионе необходимость объединения большинства данных по двум видам песчанок, именно в таком ключе провели анализ пространственной структуры их совместных поселений.
93
На участке Бармак заселенность (Z)обоими видами песчанок и каждым видом в отдельности была минимальной весной 1985 и 1987 гг. В связи с низкой численностью гребенщиковых песчанок и обнаружением их городков практически в тех же ячейках, что и полуденных, картина обобщенной пространственной структуры мало отличалась от того, что было сказано выше о полуденных песчанках. Анализ заселенности в сгруппированных ячейках показал, что все эти группы могли быть отнесены к категории «поселение». Весной 1985 г. таких групп было три: из 6 ячеек (Z = 0.47%), из 7 ячеек (Z = 0.91%) и из 12 ячеек (Z = 0.53%). Весной 1987 г. (следующая депрессия численности) не считая обособленных заселенных ячеек, были обнаружены две группы: из 12 ячеек (Z = 0.70%) и из 24 ячеек (Z = 0.63%). Средняя заселенность всех ячеек, имевших городки песчанок на этом участке весной 1985 г., составила 0.63 ± 0.06%, весной 1987 г. — 0.60 ± 0.05%. Совместные поселения двух видов песчанок меняли свою дислокацию от одной депрессии к другой, располагаясь рядом с предыдущими местами, то есть не далее чем в 1-2 км от них (рисунок 6).
Конфигурация пространственной структуры поселений песчанок обусловливается тремя процессами: размножением, смертностью и перемещениями грызунов. Среди последних играют роль только перемещения, связанные со сменой зверьками своих мест обитания (переселения, миграции). Наличие таких перемещений можно зарегистрировать, изучая изменения дислокации поселений, происходящие от осени к весне следующего года.
94

Рис. 6. Дислокация поселений полуденных и гребенщиковых песчанок в различные депрессии численности на участке «Бармак» — весной 1985 г. (А) и весной 1987 г. (Б). Ячейки, заселенные в одну из депрессий (1) и в обе (2).
I
95
В качестве наглядного примера мы выбрали осень 1984 г. и весну 1985 г. (рисунок 7). Той осенью в Бармаке было заселено 98 ячеек, весной — 38. Весной остались заселенными только 27 ячеек из 98, бывших заселенными осенью, а 11 (из 38) оказались на новых местах. Размножения в эту зиму отмечено не было, а численность песчанок на участке сократилась более чем в три раза (заселенность участка в целом составила осенью 0.55%, весной — 0.15%). Оценивая среднюю заселенность только по ячейкам, имевшим городки, численность песчанок изменилась гораздо в меньшей степени: с 0.91% до 0.63%.
Выявленные особенности динамики поселений малых песчанок были характерны и для участка в урочище Истай. Однако здесь отмечены и некоторые отличия. Во-первых, численность носителей чумы в ур. Истай была постоянно в 2-10 раз выше, чем на других участках. Во-вторых, здесь почти всегда отмечалось лишь незначительное число ячеек, не имевших городков полуденных или гребенщиковых песчанок. В связи C этим в ур. Истай не могло быть обнаружено резких передислокаций поселений зверьков, как на других участках, особенно от осени к весне.
Незначительные изменения дислокации поселений в разные депрессивные периоды все же были отмечены. Так, весной 1985 г. из 162 ячеек заселенными оказались 127. В следующий период депрессии весной 1987 г. были заселены 68 ячеек, причем повторно — 61, а 7 ячеек с городками оказались на новых местах. Размер сохраняющихся поселений в Истае также имеет отличия. Здесь даже в период глубочайшей депрессии численности весной 1987 г. отмечено цельное поселение из 48 ячеек, занимающих площадь 9600 га (рисунок 8). Средняя заселенность этого поселения полуденными и гребенщиковыми песчанками составила 0.70 ± 0.06%. Расстояние между городками в таком поселении (в случае их равномерного рассредоточения) равно 128 м, а общее их количество — около 7 тысяч. В едином поселении с таким количеством городков весной обитает около 5 тыс. песчанок, осуществляющих норовые контакты между
96

Рис. 7. Передислокация поселений полуденных и гребенщиковых песчанок от осени 1984 г. (А) к весне 1985 г. (Б) на участке «Бармак». Ячейки, заселенные осенью (1), весной (2) и в оба сезона (3).
97
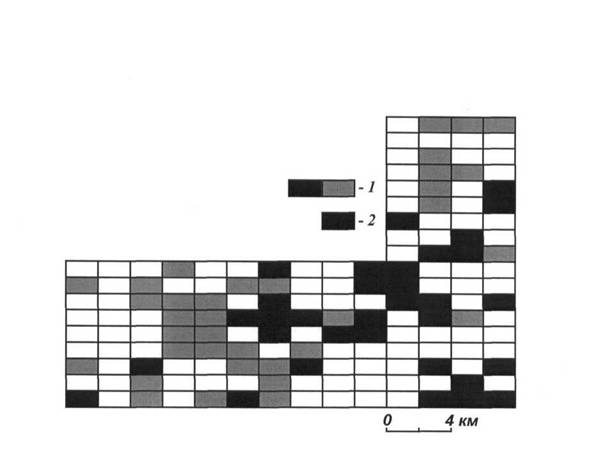
Рис. 8. «Ядро» поселений полуденных и гребенщиковых песчанок в одну из глубочайших депрессий численности на участке «Истай». Ячейки, заселенные весной 1987 г. (1) и те из них, которые были заселены в течении всех 9 сезонов картографирования 1983-1987 гг. (2).
98
собой. Кроме того, 35 ячеек на этом участке постоянно имели поселения песчанок, из них 23 — в пределах крупнейшего поселения весны 1987 г.
В итоге можно сделать вывод, что поселения полуденных и гребенщиковых песчанок в Волго-Уральских песках весьма динамичны во времени и пространстве. При наличии побуждающих мотивов зверьки способны менять дислокацию своих поселений на довольно большое расстояние за достаточно короткое время (полгода). В период депрессии численности, как правило, сохраняются небольшие разрозненные . поселения песчанок. В следующий период депрессии дислокация таких поселений на большей части песков более чем наполовину, а иногда почти полностью не совпадает с предыдущей. Однако существуют территории, где сравнительно плотные и обширные поселения полуденных и гребенщиковых песчанок сохраняются при любом уровне численности. Пространственная устойчивость этих поселений имеет своеобразные черты и требует специального рассмотрения.
При проведении сопряженного зоологического и ландшафтного картографирования западной части Волго-Уральских песков (около 2 млн. га) было выяснено, что полуденные песчанки в период депрессии численности предпочитают устраивать свои поселения в фации полыни песчаной прогрессирующей (крупнобугристые полузакрепленные пески), а гребенщиковые — в фации кустарников. И те, и другие песчанки «выделяли» по 60% своих популяций для заселения предпочитаемых ими фаций (Кузнецов с соавт., 1990). Заметим, что обширные урочища с зарослями джузгуна часто имеют признаки полузакрепленных песков и охотно заселяются обоими видами песчанок.
На участке Истай урочища с абсолютным преобладанием зарослей джузгуна занимают более половины его площади. Такие урочища наиболее предпочитаются песчанками и довольно многочисленны в центре ВолгоУральских песков в Кошалакской местности (Ралль, 1937; Матросов с соавт., 1987; Кузнецов с соавт., 1989; 1990; 1991; Матросов, 1992;
99
Матросов, Кузнецов, Ерофеев, 1996). В пределах этих урочищ отмечено поселение малых песчанок, не терявшее своей целостности, крупных размеров и оптимальной плотности зверьков даже в периоды глубочайших депрессий численности. При этом замечено, что его дислокация в два изученных нами периода депрессии совпадала не менее чем на 8 тыс. га (83% от наименьшей зарегистрированной нами площади поселения).
По признаку устойчивости подобные поселения могут быть охарактеризованы как «постоянно существующие скопления песчанок (ядра)» (Эйгелис, 1980). Из этой же работы следует, что кроме критерия постоянства во времени и пространстве, ядро поселений должно характеризоваться определенным уровнем нижнего порога численности животных, обитающих в нем, или иными словами — размером поселения, претендующего на придание ему статуса «ядра». Для закавказских видов песчанок Ю.К.Эйгелис (1980) определяет минимальную численность зверьков, населяющих ядро, не менее 1 тыс. особей. В нашем случае при минимальной заселенности внутри поселения, равной 0.29% (см. табл. 28), при которой обеспечиваются норовые контакты между животными, указанное число песчанок (1 тыс. особей) займет не менее 3 тыс. га территории. Следовательно, по всем указанным параметрам поселение полуденных и гребенщиковых песчанок на участке Истай (5 тыс. особей при заселенности 0.7%) может считаться ядром.
Это пространственно-структурное образование обладает некоторыми специфическими чертами. Во-первых, сохранение во время сильных депрессий численности таких относительно крупных объединений (ассоциаций) обитаемых нор характерно только для обширных ландшафтных фаций «заросли кустов джузгуна». Обширность этих фаций (тысячи и десятки тысяч гектар) автоматически возводит их в ранг урочищ. Во-вторых, оставаясь в границах таких урочищ (при очередной депрессии численности), поселение может частично изменить дислокацию. Ранее, свойство постепенной передислокации устойчивых поселений послужило
IOO
для нас основанием не применять к ним термин «ядро» (Кузнецов с соавт., 1991). Однако мы вправе именовать крупное скопление нор и обитающих в них животных ядром в любом случае, даже при его незначительном и медленном перемещении по территории. Поскольку «новое» ядро образовано потомками особей, населявших подобное ядро в предыдущий неблагоприятный период, то именно наличие этой «наследственности» служит главным критерием его постоянства. Постоянно существующие крупные скопления грызунов фактически являются популяционными ядрами, а территория, где эти ядра (одно или несколько) циркулируют — «экологическим центром» популяции. Число экологических центров ареала отдельной популяции или вида в целом не ограничивается. Возможно, что у некоторых популяций экологические центры могут отсутствовать.
Таким образом, в Волго-Уральских песках имеются ядра (экологические центры) популяций полуденных и гребенщиковых песчанок, совпадающие с обширными урочищами бугристых закрепленных песков с преобладанием песчанополынно-джузгуновых ассоциаций (Матросов с соавт., 1987; Кузнецов с соавт., 1990). В пределах этих урочищ постоянно существуют популяционные ядра, дислокация, площадь и конфигурация которых в каждую следующую депрессию численности могут не полностью совпадать с предыдущей (Кузнецов с соавт., 1989; 1991). Очевидно, что популяционные ядра имеют особое значение для динамики, как самих популяций грызунов, так и сопутствующих им компонентов экосистем: эктопаразитов, возбудителей заболеваний и т. д. Следовательно, ландшафтная структура ВолгоУральского песчаного очага чумы играет формирующую роль в его эпизоотологической дифференциации.
IOl
3.3.2.
Еще по теме Ландшафтная приуроченность элементов пространственной структуры популяций песчанок в ВолгоУральском песчаном очаге чумы.:
- Структура Волго-Уральского песчаного очага чумы.
- Перемещения полуденных и гребенщиковых песчанок в Волго-Уральских песках.
- Волго-Уральский песчаный очаг(16)
- Структура Прикаспийского песчаного очага чумы
- Оценка биоценотической и пространственной структуры очагов чумы
- Форезия блох в Волго-Уральских песках
- Естественная и антропогенная трансформация Прикаспийского песчаного очага чумы.
- Оценка активности природных очагов чумы — основа их пространственной дифференциации.
- Структура поселений и численность носителей и переносчиков возбудителя чумы на плато Тайнгуен
- 12.2.5. Возможная величина и структура санитарных потерь в очаге ядерного поражения
- Оценка трансмиссивного пути передачи возбудителя чумы и структура коэффициента трансмиссивности
- Прикаспийский песчаный очаг (43).
- Популяции и выборки
- 40. Нарушение генетического равновесия в популяции.
- 4.2. Функциональная гетерогенность популяций эндотелия