Динамика автолиза дрожжей
Определив преимущества СНЖК как индуктора автолиза микроорганизмов, вызывающих нарушения структурной организации и функциональной активности клеточных мембран, остановимся на выяснении условий оптимизации саморазрушения клеток, что является вторым этапом в разработке технологии.
Как было отмечено выше, о начавшемся автолизе свидетельствуют ингибирование энергодающих процессов, падение числа КОЕ и увеличение в инкубационной среде количества низкомолекулярных соединений. Из этих параметров наиболее практически важен и при этом прост в методическом исполнении и легко стандартизуем последний. Поэтому разработку режимов автолиза целесообразно вести попоказателям накопления в инкубационной среде продуктов гидролиза клеточных биополимеров.
На рис. 6 приведены данные о влиянии температуры и pH инкубационной среды на протеолитическую активность автолитического комплекса дрожжей. Об активности протеаз можно судить по образованию в результате гидролиза белков полипептидов с числом аминокислотных остатков не менее 4 (биуретовая реакция), а так же по накоплению в среде свободных аминокислот, определяемых формольным титрованием.
Первая отмеченная закономерность — несовпадение зависимости ингибирования дыхания и разрушения клеток от pH среды.
Если ингибирование дыхания и падение числа КОЕ максимальны при pH 3-4 (рис. 3), то преимущественное высвобождение из клеток продуктов гидролиза белка наблюдается при pH 6-8 (рис. 6). Не менее важно, что максимальное ингибирование
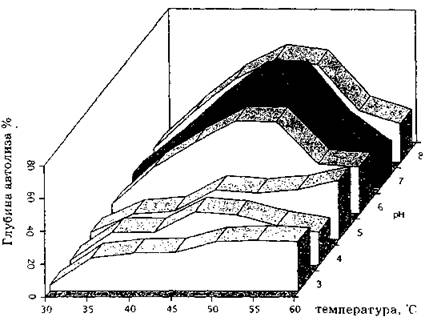
Рис. 6. Зависимость глубины автолиза клеток Ft. aurantiaca от условий ведения процесса Индуктор препарат СНЖК, 80 мкг мл; время ведения процесса - 4 ч; глубина автолиза количество растворимого белка в инкубационной сребе, от общего содержания белка в нативных клетках
дыхания (рис.
1) и критическое падение КОЕ (до 0,01-0,005%) совпадают по времени и отмен; ются через 10-15 мин после внесения индуктора в подкисленную клеточную суспензию. За эт< период времени практически не регистрируется выход из клеток низкомолекулярных соедин< ний: клетки остаются интактными, но уже становятся нежизнеспособными. Таким образом, к; тастрофическое снижение числа КОЕ в клеточной суспензии является первым признаком автг лиза. Так как определение количества КОЕ — процедура длительная, для быстрого определени нежизнеспособности можно использовать витальное окрашивание, хорошо дифференцирующе живые и неживые клетки.Приведенные результаты еще раз акцентируют внимание на том факте, что процесс индукци автолиза и собственно автолитическое разрушение структур различаются принципиально как п цели, так и по условиям достижения этой цели. При разработке технологических режимов автг лиза микроорганизмов (дрожжей) процесс следует индуцировать внесением СНЖК при подкиг лении среды до значений pH < 4,5, а саму деструкцию клеток вести в близкой к нейтральной ил слабощелочной областях pH (до 6,5-7,8), оптимальных для клеточных протеиназ. Зависимост гидролиза биополимеров от температуры стандартна для кинетики ферментативных реакци (рис. 6), и скорость гидролиза возрастает с увеличением температуры. Следует отметить, что ш вышение температуры выше 60 "С всегда приводит к снижению скорости автолиза дрожже вплоть до его прекращения, что объясняется инактивацией дрожжевых протеаз, вследствие на рушения интактности лизосом и образования неактивных комплексов лизосомальных протеаз ингибиторами, находящимися в цитозоле.
Проанализируем вышеизложенные данные. Еще раз напомним, что автолитические процес сы — это постоянно присутствующая активность биохимических гидролитических процессов определяющих физиологию роста и развития любой клетки: от оборота клеточного материал и субклеточных структур до роста клетки и ее разделения (септации, почкования, перетяжки дробления, апикального роста и т.д.).
При этом скорости процессов деградации и синтеза моле кул биополимеров сбалансированы и величина этого баланса определяет уровень метаболичс ской активности клетки и, соответственно, стадию развития культуры, т.е. скорость ее рость старения и отмирания клеток.Дисбаланс синтетических и автолитических процессов, естественно возникающий в клетка стареющей культуры или являющийся следствием внешних воздействий (индукции), в польз; автолитических, является причиной необратимого разрушения клеточных структур и, в перву? очередь, клеточной оболочки, что приводит к гибели клеток.
Поэтому для разработки технологий автолиза существенное значение имеют, с одной сторс ны, знания об ультраструктурной организации эукариотной клетки, ее органеллах, локализь. ции ферментов и их ингибиторов; с другой — информация о природе гидролаз, включающих ка: истинные автолизины, разрушающие материал клеточной стенки, и липазы, гидролизующи мембранные липиды, так и ферменты протеолитического комплекса, являющиеся ключевым, и для осуществления физиологических автолитических процессов, и для автолиза как процесс гибели клетки.
Табл. 3. Влияние подкисления среды при индукции автолиза на интенсивность саморастворения клеток дрожжей R.aurantiaca (12% МСК)
| pH | Продолжительность автолиза, ч | Концентрация СНЖК, нМ/мл | Содержание растворимого белка, мг/г МСК |
| pH 6,8-7,0 | 6 | 600 | 201,3 |
| сразу pH 3,5 - 1 мин.затем pH 6,8-7,0 | 2 Н | 200 | 120,3 |
| 4 | 200 | 168,0 | |
| 6 | 200 | 198,9 | |
| pH 3,5 - 15 мин, затем pH 6,8 | 2 | 200 | 115,6 |
| 4 | 200 | 170,4 | |
| ; 6 | 200 | 207,7 | |
| pH 3,5 - 30 мин, затем pH 6,8 | 2 | 150 | 95,4 |
| і 4 | 150 | 180,0 | |
| 6 | 150 | 190,6 |
Табл. 4.
Автолиз клеток R. aurantiaca, индуцированный внесением предварительно полученного автолизата| Количество СНЖК в «затравке», нМ/мл | Объем внесенной «затравки», % | Добавление СНЖК к общему объему, нМ/мл | Общее количество С.НЖК в пробе, нМ/мл | Содержание остаточного белка в клетках дрожжей мг/г МСК |
| 600 | 50 | 600 | 785 | 16,39 ± 3,04 |
| 600 | 50 | - | 200 | 43,25 ± 4,12 |
| 1200 | 50 | 600 | 900 | 15,33 ± 4,78 |
| 1200 | 50 | - | 400 | 42,46 ± 2,74 |
| 1800 | 50 | 600 | 1200 | 19,72 ± 4,78 |
| 1800 | 50 | - | 600 | 33,48 ± 7,78 |
| Контроль (нативные клетки) | - | - | 180 ± 5,60 |
Из протеиназ дрожжевых клеток особое внимание привлекает Ysc А, локализованная в активной форме в лизосомах, по выходе из которых она связывается с ингибитором пепстатином, находящимся в цитозоле, и переходит в неактивный комплекс, который разрушается с высвобождением активного энзима либо при низких pH, либо вследствие отщепления ингибитора проте- иназой Ysc В, локализованной также в лизосомах. Этим объясняется несоответствие в рН-опти- мумах для индукции автолиза (создания при низких значениях pH необратимого дисбаланса анаболических и катаболических процессов за счет активации протеиназы Ysc А) и для реакций последующего разрушения биополимеров, в первую очередь, белков, с участием основных дрожжевых протеиназ, имеющих pH-оптимум в нейтрально-щелочной области.
Другой процесс, имеющий важное значение для индукции автолиза, — это нарушение структурного состояния и функциональной активности ЦПМ, а также мембран внутриклеточных органелл и ЭПР. Изменение жидкокристаллического состояния мембранных липидов в сторону необратимого увеличения их текучести приводит к нарушению барьерных и других функций мембран, вследствие дезорганизации липидной стромы мембраны и нарушений липид-белковых взаимодействий интегрированных и ассоциированных с ней ферментов. Поэтому супраопти- мальная концентрация внеклеточных СНЖК, вызывающая дестабилизацию мембран, является второй самостоятельной причиной индукции автолиза. Закисление инкубационной среды, способствующее более эффективному включению в мембраны протонированных молекул СНЖК, позволяет снизить нормы их расхода для индукции автолиза (табл. 3).
Как следует из приведенных данных, автолиз, индуцированный внесением СНЖК при одновременном подкислении инкубационной среды, и создание затем оптимальных условий протекания собственно автолитической деструкции клеток позволяют проводить равно глубокий гидролиз внутриклеточных биополимеров при использовании индуктора в значительно меньшей концентрации (150-200 нМ вместо 600 нМ) и сокращают время автолиза до 4 ч вместо 6 ч.
От среды роста, т.е. стресс,вызванный высокой клеточной плотностью, и лишение клеток источников энергии, что,согласно постулату Вэлча,индуцирует автолитическую деструкцию клеток.
И, наконец, последняя причина развивающихся литических процессов — собственно образующийся автолизат, который может служить затравкой и применяться как индуктор, благодаря действию содержащихся в нем экзогенных (вышедших из клеток в инкубационную среду) гидролаз. Дополнительно к гидролазам в автолизате-затравке действуют и вышедшие из автолизирующихся клеток СНЖК.
В качестве примера использования самой автолизированной суспензии для индукции автолиза рассмотрим процесс гидролитического разрушения биомассы «ожиренных» (выращенных в режиме преимущественного накопления липидов) дрожжей R.
aurantiaca, выращенных на солевой среде с этанолом (табл. 4). Автолизат-затравку получали в результате индуцированного СНЖК автолиза клеток этих же дрожжей фазы линейного роста при pH 7,5 в течение 8 ч. Полученный автолизат использовали в качестве индуктора, добавляя его в количестве 50% (об./об.) к основной биомассе дрожжей, подщелоченной до pH 6,8-7,2. Помимо этого в некоторые пробы дополнительно вносили СНЖК в количестве 350 нМ/мл. Предварительно определили, что в нативных клетках дрожжей содержалось белка около 180 мг/г МСК. Процесс автолиза (табл. 4) позволил реализоватьв инкубационную среду до 40-45% белка клетки в виде низкомолекулярных соединений, а в ваі антах внесения дополнительного количества индуктора — углубить протеолиз с выходом иепти; до 60-75% от общего белка клеток.
Прием использования отъема автолизата как затравки для индукции автолиза особенно целе< образен при разработке технологии автолиза в непрерывном режиме для получения глубоко гид] лизованных продуктов, обогащенных в основном свободными аминокислотами.
Анализ данных, приведенных в этом разделе, обосновывает вывод о двустадийности как ин; цированного, так и спонтанного автолиза микроорганизмов, где первой стадией является инду ция, регистрируемая по ингибированию энергодающих процессов и падению числа жизнеспосс ных клеток, а второй — собственно автолитическое разрушение клеток, определяемое по выхо в инкубационную среду продуктов деградации биополимеров. Подчеркнем, что эти стадии разл чаются принципиально как по природе и итогу процесса, так и по условиям, оптимальным для дс тижения этого итога.
К параметрам, которые необходимо учитывать на стадии индукции относятся: pH инкубационн< среды; плотность автолизируемой клеточной суспензии; концентрация индуктора и способ его внес ния. Оптимизация этого этапа позволяет существенно (в 2 и более раз) сократить расход индукто; (табл. 3), а также влияет на скорость и глубину автолиза на стадии деградации клеток.
3.4.
Еще по теме Динамика автолиза дрожжей:
- Автолиз дрожжей в периодическом режиме
- Автолиз дрожжей в непрерывном режиме
- Технология автолиза дрожжей
- Разработка технологии автолиза дрожжей
- 3.2.2. Влияние иммобилизованных дрожжей на динамику физико-химических показателей кюве
- Регистрируемые параметры автолиза
- Автолиз и его индукция
- Индукторы автолиза
- Практическое применение автолизатов дрожжей
- Мембранотропные соединения как индукторы автолиза
- Авто лизаты кормовых дрожжей
- Выбор штамма дрожжей
- Экстракты автолизированных дрожжей
- 3.3. Получение красных игристых вин на основе использования иммобилизованных дрожжей
- Технология получения белковых селенобогащенных БАД на основе дрожжей
- 3.2.1 Особенности вторичного брожения с использованием дрожжей, иммобилизованных в геле альгината натрия из водоросли рода Cistozeira.
- Получение селенобогащенной биомассы дрожжей