Регистрируемые параметры автолиза
Первой характеристикой необратимости автолитической деградации и гибели клеток является потеря ими жизнеспособности, что определяется как резкое снижение численности колониеобразующих единиц (КОЕ) при высеве проб автолизирующейся суспензии на плотные питательные среды (в случае автолиза дрожжей — на сусло-агар).
О дальнейших изменениях, происходящих в процессе автолиза, можно судить по изменению реологических свойств клеточной суспензии. На начальных стадиях автолиза происходят прежде всего изменения, связанные с разрушением клеточных оболочек: необратимая структурная дезорганизация ЦПМ и потеря ею барьерной функции; падение тургора клетки, вследствие диффузионного выхода из нее ионов и ряда низкомолекулярных соединений; уменьшение за счет этого размеров клетки. Эти события обусловливают изменения вязкости клеточной суспензии.
В следующей фазе происходит гидролитическое расщепление клеточных биополимеров, разрушение внутриклеточных структур и высвобождение продуктов гидролиза в инкубационную среду. Клеточная стенка во время автолиза эндотипа подвергается определенным модификациям, и, хотя она разрыхляется и частично гидролизуется, целостность ее сохраняется. У микроорганизмов с экзотипом автолиза клеточная стенка разрушается полностью. Подчеркнем, что необходимым условием автолитической деструкции клетки является полное разрушение цитоплазматической мембраны. У дрожжей автолиз протекает по эндотипу и начинается с разрушения ЦПМ. У некоторых грибов, обладающих высокой активностью эндо- и экзоглюканаз, имеет место экзотип автолиза и наблюдается полное разрушение не только мембраны, но и клеточной стенки.
О развитии автолитических процессов часто судят по падению оптической плотности клеточных суспензий. Однако более корректно характеризовать автолиз по биохимическим показателям: высвобождению в инкубационную среду продуктов деградации клеточных биополимеров и снижению их исходного содержания в клетке, например, уменьшению количества внутриклеточного белка параллельно с увеличением количества продуктов его гидролиза в инкубационной среде.
Характеристиками автолиза служат интенсивность процесса и глубина распада клеточных структур.
Интенсивность автолиза определяется скоростью реализации продуктов гидролиза клеточных биополимеров и как суммарный ферментативный процесс зависит от:
• количества гидролаз;
• активности гидролаз;
• физико-химических условий среды;
• субстратной устойчивости клеточных компонентов к действию ферментов автолитического комплекса.
Глубина автолиза измеряется соотношением общего количества имеющихся в клетке до автолиза биополимеров (белков, полисахаридов, нуклеиновых кислот) и их гидролизовавшейся частью. Глубина и скорость автолиза зависят от физиологического возраста клеток. Клетки экспоненциально растущей культуры автолизируются быстрее и в большей степени, чем клетки стационарные, полного саморастворения которых, как правило, добиться не удается. Это объясняется тем, что наиболее интенсивно ферменты автолитического комплекса синтезируются в клетках экспоненциальной фазы роста, где они принимают участие в важнейших физиологических процессах, таких как рост и деление клеток. Зависимость автолиза от возраста связана не только с количеством имеющихся в клетке ферментов автолитического комплекса, но и с их активностью, что обусловлено изменением конформации ферментов и физико-химических условий в стареющих клетках. Например, маннаназы в стационарных клетках дрожжей менее активны, чем в экспоненциальных.
При разработке технологий автолиза важно учитывать последовательность событий, приводящих к деструкции микробных клеток. Прежде всего, это относится к разграничению этапов индукции автолиза и собственно автолитического разрушения клеточных биополимеров, различающихся как по целевой направленности происходящих на этих этапах процессов, так и по условиям, определяющим их развитие. Индукция автолиза предусматривает создание дисбаланса между процессами синтеза и деградации клеточных структур. Ее результативность определяется харак-
Табл. 1. Влияние коммерческих препаратов жирных кислот на дыхание интактных клеток фазы линейного роста дрожжей R.aurantlaca
| Жирная кислота | Остаточное дыхание клеток (% к контролю) | ||||
| при добавлении препарата в концентрации | 800 нМ/мл | ||||
| 25 нМ/мл | 200 нМ/мл | 300 нМ/мл | 600 нМ/мл | ||
| Пальмитиновая (С 16:0) | 106,0 | 100,5 | 103,0 | 93,5 | 84,2 |
| Стеариновая (С 18:0) | 102,0 | 112,0 | 95,0 | 84,0 | 78,1 |
| Олеиновая (С 18:1) | 142,5 | 43,0 | 27,0 | 25,5 | 20,0 |
| Линолевая (С 18:2) | 95,0 | 40,0 | 25,0 | 22,0 | 20,0 |
тером выбранного индуктора и физико-химическими условиями, оптимальными для лишения клетки энергии или нарушения структуры клеточной оболочки.
Интенсивность автолиза и глубина разрушения клеточных биополимеров зависят от количества ферментов автолитического комплекса и условий, оптимальных для проявления их активности.Для получения необходимой информации об эффективности этапов автолиза целесообразно проводить исследования временных характеристик (динамики) клеточной деструкции, которые дают возможность определить сроки интенсивного развития процесса, а также временные интервалы для корректировки физико-химических параметров среды для поддержания высокой скорости гидролиза клеточных биополимеров.
3.2. Выбор индуктора
Выбор эффективного индуктора является первой технологической задачей. В этом отношении предпочтение следует отдать СНЖК, т.к., во-первых, они являются естественным компонентом клетки и поэтому нет необходимости удалять их из полученного продукта — автолизата; во-вторых, они производятся из возобновляемого сырья и потому достаточно дешевы и доступны.
Хорошим биологическим тестом для подбора индуктора является определения его влияния на дыхание клеток, так как этот показатель количественно отражает изменения функциональной активности мембран, обусловленные нарушениями их структурной организации, что, как было сказано выше, является причиной автолиза. Эндогенное дыхание клеток измеряют полярографически в кислородной ячейке конструкции Шольца-Островского.
Биологическое тестирование коммерческих препаратов индивидуальных жирных кислот показывает, что в основном за физиологическое действие — ингибирование дыхания и индукцию авто-
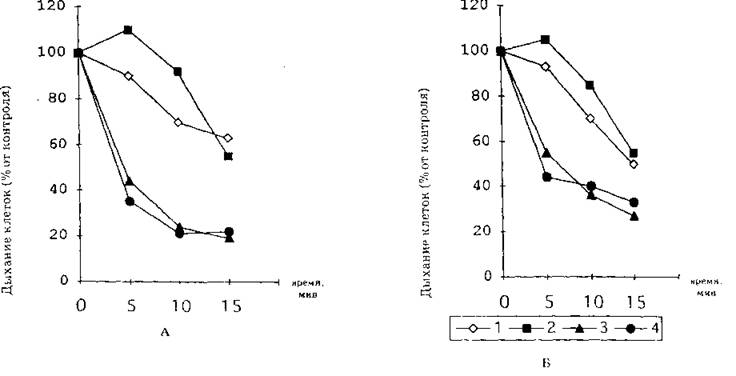
Рис. 1. Влияние пальмитиновой (I ), стеариновой (2), олеиновой (3), линолевой ( 4 ) кислот в концентрации 600 нМ/мл на дыхание клеток fLauratiaca (Л) и S.serei’isi'uc ( Ц /
Табл. 2. Индукция автолиза суспензии дрожжей S.cerevisiae (МСК 12%) коммерческими препаратами жирных кислот
| Жирная кислота | Концентрация индуктора, мас.% | Реализованный белок, % |
| Пальмит-олеиновая С16:1 | 1:1250 | 39,3 |
| Олеиновая С18:1 | 1:1000 | 26,3 |
| Линолевая С18:2 | 1:1666 | 32,1 |
| Линолеяовая С18:3 | 1:2000 | 49,8 |
лиза клеток — ответственны ненасыщенные жирные кислоты (табл. 1), хотя все испытанные жир- ные кислоты в концентрациях более 300 нМ/мл вызывают ингибирование эндогенного дыхания клеток дрожжей R.aurantiaca.
Высокие концентрации СНЖК (200-800 нМ/мл), приводя к сверхкритическому увеличению жидкостности клеточных мембран, вызывают ингибирование дыхания клеток любой фазы роста. Повышение концентрации до 1000 нМ/мл и более не приводит к ускорению или увеличению степени ингибирования дыхания клеток, видимо, из-за эффекта мицеллообразования вокруг клетки за счет избытка жирных кислот.
Эффект ингибирования дыхания развивается во времени (рис. 1). Степень ингибирования дыхания клеток дрожжей под воздействием отдельных жирных кислот различается значительно: через 10 мин после внесения в клеточную суспензию отмечается подавление дыхания дрожжей олеиновой кислотой на 62%, линолевой — на 61%, пальмитиновой — на 29%, стеариновой — на 18%. Коммерческие препараты ненасыщенных жирных кислот в концентрациях, подавляющих дыхание клеток дрожжей, эффективно индуцируют автолиз (табл. 2).
Различия в дестабилизирующем мембраны действии различных жирных кислот (изменение фазового состояния мембран) связаны с разницей в температурах их плавления — более низких у ненасыщенных ЖК: стеариновая кислота — 69,6 ”С, пальмитиновая — 63,1 ’С, олеиновая — 13,5 “С, линолевая — минус 5 °С. Кроме того, повышение степени ненасыщенности мембранных липидов коррелирует с увеличением проницаемости мембран, а повышение концентрации СНЖК вызывает разобщение дыхания и окислительного фосфорилирования. Суммарные модификационные изменения структурных и функциональных свойств мембран, вызванные дестабилизирующим действием ненасыщенных ЖК, играют драматическую роль в развитии патологических процессов, одним из которых является автолиз клеток, завершающийся их гибелью.
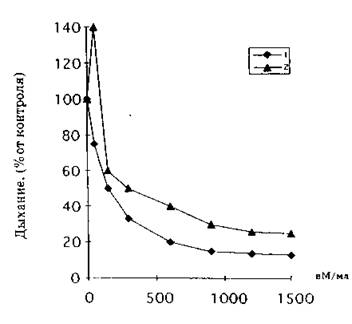
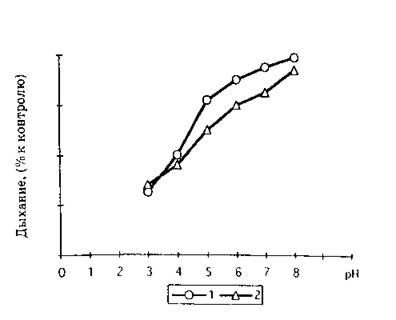
Рис. 3. Влияние препарата СНЖК на эндогенное дыхание клеток R. auratiaca (1 ) и S. cerevisiae (2) линейной фазы роста в зависимости от pH среды (0.15 .М фосфат цитратный буфер, концентрация СНЖК 30 мкг/мл )
Рис.
2. Влияние олеиновой кислоты на эндогенное дыхание клеток R. av’-atiaca. взятых с экспоненциальной (1) и середины линейной (2) фаз роста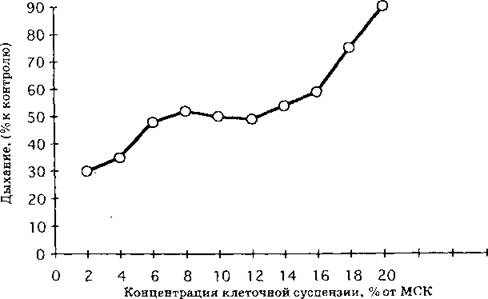
Рис. 4. Влияние препарата СНЖК на эндогенное дыхание клеток R. aurati- аса линейной фазы роста в зависимости от концентрации клеточной сус пензии (концентрация СНЖК 80 мкг!мл ) )
Биологические тесты влияния на дыхание клеток целесообразно использовать и для определения эффективности действия и подбора доз индуктора. Сравнительный анализ действия коммерческих препаратов ненасыщенных жирных кислот показывает общие закономерности их эффектов. Для всех СНЖК степень ингибирования дыхания клеток повышается при: увеличении концентрации вносимых липидов (рис. 2), снижении pH инкубационной среды (рис. 3), уменьшении количества биологического материала в ячейке (рис. 4), а также развивается во времени (рис. 1) и зависит от физиологического возраста клеток (рис. 2, 5).
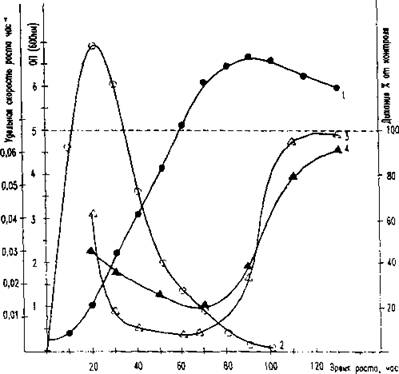
Рис. 5. влияние препарата СНЖК и олеиновой кислоты па дыхание клеток периодичес кой культуры fi.au rat iaca 1 - кривая роста, ОН (600 нм); 2 - удельная скорость роста дрожжей (р), ч '}; 3 - дыхание клеток при добавлении препарата СНЖК (80 мкг/мл клеточной суспензии );
4 — дыхание клеток при внесении олеиновой кислоты (600 нМ/мл клеточной сиспен .ши )
Обратим внимание, что на рис. 2 и 5, характеризующих влияние свободных жирных кислот на дыхание клеток различного физиологического возраста, имеется существенное различие в их действии на клетки экспоненциальной, линейной и стационарной фаз роста. Во-первых, только у клеток линейной фазы роста или более старых отмечается стимуляция дыхания при низких концентрациях СНЖК, в то время как у экспоненциально растущих клеток эти дозы вызывают ингибирование дыхания и индуцируют автолиз. Во-вторых, степень ингибирующего действия снижается по мере старения микробной культуры.
Эффект стимуляции дыхания объясняется двояко: протонофорным действием СНЖК, которые, в отличие от классических протонофоров, обнаруживают это свойство, когда, пройдя через митохондриальную мембрану, обменивают протон карбоксильной группы на внутриклеточный калий, а также способны в значительной степени снижать электрическое сопротивление мембран, что совокупно приводит
к нарушению электрического трансмембранного потенциала и компенсаторному увеличению дыхания при разобщении с биоэнергетическими процессами. Другое объяснение относится к способности СНЖК повышать текучесть мембран и, следовательно, активность интегрированных в них ферментов.
Липиды мембран клеток экспоненциальной фазы роста в наибольшей степени обогащены ненасыщенными ЖК, т. е. обладают мембраной, текучесть которой приближается к максимально возможной, что и определяет максимальную физиологическую активность клеток этой фазы роста, но также и максимальную их чувствительность к индукторам автолиза, поскольку даже незначительное повышение текучести липидов в их мембранах является сверхкритическим.
Отсюда следует важный вывод, что если наибольшей чувствительностью к действию СНЖК обладают клетки экспоненциальной и линейной фаз роста, то в работе с непрерывными культурами необходимые дозы индуктора автолиза при высоких скоростях протока должны быть значительно меньше, чем требующиеся для автолиза клеток стационарных фаз роста периодических культур. Этот вопрос будет рассмотрен далее более подробно.
Нужно отметить, что, хотя в тестах на дыхание клеток дрожжей определяется действие СНЖК на функциональную активность митохондриальных мембран, но и ЦПМ дрожжевой клетки, и мембраны ЭПР, и множество других клеточных структур, оформленных при участии мембран (в том числе лизосомы), также являются мишенью для действия свободных жирных кислот.
При разрушении мембран лизосом под действием СНЖК начинается неконтролируемое расщепление макромолекул и клеточных структур под действием лизосомальных гидролаз, которое заканчивается гибелью клетки.
Естественно, что, обладая протонофорным действием, СНЖК проявляют большую активность в подкисленной среде, находясь в протонированном состоянии (рис. 3), что может быть использовано для снижения норм их расхода при индукции автолиза. Также следует обратить внимание на зависимость ингибирования дыхания жирными кислотами от плотности клеточной суспензии (рис. 4), что также должно учитываться при расчетах эффективных доз индуктора и будет более подробно рассмотрено ниже.
Таким образом повышение уровня СНЖК в клеточной суспензии влияет на энергетическую функцию (дыхание) и фазовое состояние мембран вплоть до их дестабилизации, что является побудительной причиной автолиза клеток микроорганизмов различных таксономических групп, как про-, так и эукариот.
Результаты, свидетельствующие о видонеспецифичности действия СНЖК как эффективного индуктора автолиза; промышленном производстве препаратов СНЖК; нетоксичности этих соединений, благодаря чему их не требуется удалять из конечного продукта, — позволяют рекомендовать СНЖК для технологий автолиза микроорганизмов в качестве универсального индуктора.
3.3.
Еще по теме Регистрируемые параметры автолиза:
- Анализ вариабельности регистрируемых параметров в оптической неинвазивной диагностике
- Динамика автолиза дрожжей
- Автолиз и его индукция
- Индукторы автолиза
- Автолиз дрожжей в периодическом режиме
- Автолиз дрожжей в непрерывном режиме
- Мембранотропные соединения как индукторы автолиза
- Технология автолиза дрожжей
- Разработка технологии автолиза дрожжей
- Задание, управление и контроль параметров
- Измерение параметров жестких контактных линз
- Параметры АР
- Параметры центральной гемодинамики на фоне низко- и высокосолевой диеты
- Оценка влияния холодовой прессорной пробы на комплексные параметры микроциркуляторного русла