сЯКНБХЪ ПЮЯОПНЯРПЮМЕМХЪ ЩОХГННРХВЕЯЙНЦН ОПНЖЕЯЯЮ
б ТНПЛСКЕ ЙНЩТТХЖХЕМРЮ РПЮМЯЛХЯЯХБМНЯРХ НРЯСРЯРБСЧР МЕЙНРНПШЕ УПНМНЛЕРПХВЕЯЙХЕ ОЮПЮЛЕРПШ ОПНЖЕЯЯЮ ОЕПЕДЮВХ ХМТЕЙЖХХ, ЙНРНПШЕ МЕ НЙЮГШБЮЧР БКХЪМХЪ МЮ ЙНКХВЕЯРБН ГЮПЮФЮЧЫХУЯЪ НАЗЕЙРНБ, МН БЮФМШ ДКЪ НАЫЕИ УЮПЮЙРЕПХЯРХЙХ ЩОХГННРХВЕЯЙНЦН ОПНЖЕЯЯЮ.
й РЮЙХЛ ОЮПЮЛЕРПЮЛ НРМНЯЪРЯЪ: БПЕЛЪ, ОПНУНДЪЫЕЕ Я ЛНЛЕМРЮ НЙНМВЮМХЪ ЙНПЛКЕМХЪ АКНУХ МЮ ГБЕПЭЙЕ Я АЮЙРЕПХЕЛХЕИ ДН НАПЮГНБЮМХЪ С МЕЕ АКНЙЮ ОПЕДФЕКСДЙЮ, Ю РЮЙФЕ БПЕЛЪ, ОПНУНДЪЫЕЕ Я ЛНЛЕМРЮ ГЮПЮФЮЧЫЕЦН СЙСЯЮ ЦПШГСМЮ АКНЙХПНБЮММНИ АКНУНИ ДН МЮВЮКЮ С МЕЦН АЮЙРЕПХЕЛХХ. рЮ ХКХ ХМЮЪ ОПНДНКФХРЕКЭМНЯРЭ ЩРХУ ОЕПХНДНБ ЯЙПШРНЦН ПЮГБХРХЪ ЛХЙПНАМНИ ОНОСКЪЖХХ Б НПЦЮМХГЛЕ МНЯХРЕКЕИ Х ОЕПЕМНЯВХЙНБ ХЛЕЕР ГМЮВЕМХЕ ЙЮЙ ДКЪ РЕППХРНПХЮКЭМНЦН ОЕПЕЛЕЫЕМХЪ ХМТЕЙЖХХ Б ГЮПЮФЕММШУ НАЗЕЙРЮУ, РЮЙ Х ДКЪ СВЕРЮ НАЫЕИ ДКХРЕКЭМНЯРХ (Х ЯЙНПНЯРХ) ОПНЖЕЯЯЮ (яНКДЮРЙХМ, 1963; 1968; нЯРПНБЯЙХИ, яНКДЮРЙХМ, 1965; яНКДЮРЙХМ, тЕМЧЙ, 1968).хГБЕЯРМН, ВРН ЯПНЙХ НАПЮГНБЮМХЪ ВСЛМНЦН АКНЙЮ ОПЕДФЕКСДЙЮ С АНКЭЬХМЯРБЮ БХДНБ АКНУ СЙКЮДШБЮЧРЯЪ Б 3-4 МЕДЕКХ Б ЯПЕДМЕЛ Х ПЕДЙН АШБЮЧР ЛЕМЭЬЕ 6 ДМЕИ. бЯЕ ЩРН БПЕЛЪ ГЮПЮФЕММЮЪ АКНУЮ БЕДЕР НАШВМШИ ╚ГДНПНБШИ╩ НАПЮГ ФХГМХ. щРН ОНДПЮГСЛЕБЮЕР ЕЕ ЯХЯРЕЛЮРХВЕЯЙХЕ МЮОЮДЕМХЪ МЮ ОПНЙНПЛХРЕКЕИ, ХУ ЯЛЕМС Х ЯБЪГЮММСЧ Я ЩРХЛ РПЮМЯОНПРХПНБЙС АКНУХ (ТНПЕГХЧ) ХГ МНПШ Б МНПС Х РЮЙ ДЮКЕЕ. б ПЕГСКЭРЮРЕ ЩРНЦН Й ЛНЛЕМРС НАПЮГНБЮМХЪ С АКНУХ АКНЙЮ ОПЕДФЕКСДЙЮ, НМЮ ЛНФЕР НЙЮГЮРЭЯЪ Б ЯНРМЪУ Х РШЯЪВЮУ ЛЕРПНБ НР ЛЕЯРЮ ЯБНЕЦН ГЮПЮФЕМХЪ. яЙЮГЮММНЕ ДХЙРСЕР МЕНАУНДХЛНЯРЭ РЕНПЕРХВЕЯЙНЦН ПЮЯЯЛНРПЕМХЪ БНОПНЯЮ Н ПЮГМНЯЕ ЩЙРНОЮПЮГХРНБ ОН ОНЯЕКЕМХЪЛ УНГЪЕБ.
нЯНАЕММНЯРЭЧ ОПЕДЯРЮБКЕММШУ Б ОНДПЮГДЕКЮУ 5.1.1. Х 5.1.3. УПНМНЦПЮТХВЕЯЙХУ ЛНДЕКЕИ ЪБКЪЕРЯЪ НЦПЮМХВЕММНЯРЭ ПЮГЛЕПНБ ХЯОНКЭГСЕЛШУ ДКЪ ЩРНЦН ЯХЯРЕЛ ХГ МНП, ЦПШГСМНБ Х АКНУ. уНРЪ АШКН БШЪЯМЕМН, ВРН ВХЯКН БЯЕУ ЩРХУ НАЗЕЙРНБ МЕ БКХЪЕР МЮ БЕПНЪРМНЯРЭ ЯЛЕМШ ОПНЙНПЛХРЕКЪ, МН ВЕЛ ЛЕМЭЬЕ ╚ОНЯЕКЕМХЕ╩, Б ЙНРНПНЛ ЖХПЙСКХПСЧР АКНУХ, РЕЛ АШЯРПЕЕ Х ВЮЫЕ ОПНХЯУНДХР БНГБПЮР ЙЮФДНЦН ОЮПЮГХРЮ МЮ АНКЕЕ
230
ПЮММЕЦН УНГЪХМЮ.
б ЛНДЕКЪУ Я ДБСЛЪ ЦПШГСМЮЛХ АКНУЮ ЛНЦКЮ ОЕПЕИРХ Я НДМНЦН МЮ ДПСЦНЦН, ГЮРЕЛ НАПЮРМН Х РЮЙ ЛМНЦН ПЮГ, МН ЙЮФДШИ РЮЙНИ ОЕПЕУНД ПЮЯЖЕМХБЮКЯЪ ЙЮЙ НВЕПЕДМЮЪ Х ╚ОНКМНОПЮБМЮЪ╩ ЯЛЕМЮ ОПНЙНПЛХРЕКЪ. оПХ РЮЙНЛ ОНКНФЕМХХ ДЕК Б ПЕЮКЭМШУ СЯКНБХЪУ ОПХПНДМШУ НВЮЦНБ ВСЛШ ЛНЦСР БНГМХЙЮРЭ ЯХРСЮЖХХ, ЙНЦДЮ БЯЕ ХКХ ЛМНЦХЕ АКНУХ, НДМХ ХГ ЙНРНПШУ ОНАШБЮБ, Ю ДПСЦХЕ, МЕ ОНАШБЮБ МЮ МНБШУ ГБЕПЭЙЮУ, БМНБЭ НЙЮГШБЮЧРЯЪ МЮ РЕУ ФЕ ЦПШГСМЮУ, МЮ ЙНРНПШУ НМХ АШКХ ГЮПЕЦХЯРПХПНБЮМШ Б ОПНЬКШИ ПЮГ. еЯКХ ОПЕДОНКНФХРЭ, ВРН МЮЛ ЯРЮКХ ХГБЕЯРМШ РНКЭЙН ЩРХ ДБЕ ПЮГДЕКЕММШЕ БПЕЛЕМЕЛ ОПХПНДМШЕ ЯХРСЮЖХХ, РН МЮ ЩРНЛ НЯМНБЮМХХ ЛНФМН АШКН АШ ЯДЕКЮРЭ НЬХАНВМШИ БШБНД Н РНЛ, ВРН АКНУХ БННАЫЕ МЕ ЛЕМЪЧР ОПНЙНПЛХРЕКЕИ. гМЮЪ НЯНАЕММНЯРХ ХГСВЮЕЛШУ ЯННАЫЕЯРБ ЦПШГСМНБ Х АКНУ Б СЯКНБХЪУ ОПХПНДМШУ НВЮЦНБ ЛНФМН ОПЕДОНКНФХРЭ, ВРН ЯХРСЮЖХХ, АКХГЙХЕ Й РНКЭЙН ВРН НОХЯЮММНИ, ЕЯКХ Х БНГЛНФМШ, РН ДНЯРЮРНВМН ПЕДЙХ. щРН ОПЕДОНКНФЕМХЕ НЯМНБЮМН МЮ РНЛ, ВРН Б ОПХПНДЕ, ЙЮЙ ОПЮБХКН, БЯРПЕВЮЧРЯЪ ОНЯЕКЕМХЪ МНЯХРЕКЕИ, ЯНЯРНЪЫХЕ ХГ ЯНРЕМ Х РШЯЪВ НЯНАЕИ, НАЗЕДХМЕММШУ РЕППХРНПХЮКЭМН Х ЙНМРЮЙРХПСЧЫХУ ЛЕФДС ЯНАНИ ЙЮЙ МЕОНЯПЕДЯРБЕММН, РЮЙ Х ОН ЖЕОНВЙЕ ВЕПЕГ МНПШ ДПСЦХУ ГБЕПЭЙНБ.юМЮКХГ ГЮЙНМНЛЕПМНЯРЕИ ЖХПЙСКЪЖХХ АКНУ Б ЛНДЕКЭМШУ ОНЯЕКЕМХЪУ, ЯНДЕПФЮЫХУ АНКЭЬНЕ ВХЯКН ЩКЕЛЕМРНБ (ДЕЯЪРЙХ Х АНКЕЕ), РЕУМХВЕЯЙХ РПСДМНБШОНКМХЛ Я ХЯОНКЭГНБЮМХЕЛ СФЕ ХГБЕЯРМШУ УПНМНЦПЮЛЛ МЮОЮДЕМХИ АКНУ. оНЩРНЛС АШКХ ЯДЕКЮМШ ЯННРБЕРЯРБСЧЫХЕ НАНАЫЕМХЪ, ОНГБНКХБЬХЕ Б НАЫХУ ВЕПРЮУ НОХЯЮРЭ ХЯЯКЕДСЕЛНЕ ЪБКЕМХЕ.
оПЕФДЕ БЯЕЦН, АШКН МЕНАУНДХЛН ЯНГДЮРЭ ХДЕЮКХГХПНБЮММСЧ ОПНЯРПЮМЯРБЕММСЧ ЛНДЕКЭ ОНЯЕКЕМХЪ ЦПШГСМНБ-МНПМХЙНБ, НАХРЮРЕКЕИ НРЙПШРШУ ЮПХДМШУ ОПНЯРПЮМЯРБ, УЮПЮЙРЕПМШУ ДКЪ ЛМНЦХУ ОПХПНДМШУ НВЮЦНБ ВСЛШ. б ЙЮВЕЯРБЕ РЮЙНИ ЛНДЕКХ АШКЮ ОПХМЪРЮ ОПЮБХКЭМЮЪ РПЕСЦНКЭМЮЪ ПЕЬЕРЙЮ, СГКЮЛХ ЙНРНПНИ ЯКСФХКХ ХМДХБХДСЮКЭМШЕ ФХКХЫЮ (МНПШ, ЦНПНДЙХ, ЙНКНМХХ Х Р. Д.) ГБЕПЭЙНБ. пЕЬЕРЙЮ ЩРНЦН РХОЮ АШКЮ БШАПЮМЮ ОНРНЛС, ВРН ДКЪ МЕЕ УЮПЮЙРЕПМЮ МЮХАНКЕЕ ОКНРМЮЪ ╚СОЮЙНБЙЮ╩ СГКНБ ОПХ РЮЙНЛ ФЕ ПЮЯЯРНЪМХХ ЛЕФДС МХЛХ, ЙЮЙ Х Б ДПСЦХУ, ЯПЮБМХБЮЕЛШУ
231
Я РПЕСЦНКЭМНИ, ПЕЬЕРЙЮУ (йСДПХМ, гЮБЮКХЬХМ, 1973; щТПНЯ, 1982; оНЯРМХЙНБ, вХЙПХГНБ, 1983).
оПЕДОНКЮЦЮЕРЯЪ, ВРН РЮЙЮЪ ЙНЛОНМНБЙЮ СГКНБ ЙЮЙ Б ОПЮБХКЭМНИ РПЕСЦНКЭМНИ ПЕЬЕРЙЕ ЪБКЪЕРЯЪ ХДЕЮКНЛ, Й ЙНРНПНЛС ЯРПЕЛЪРЯЪ ЦПШГСМШ ОПХ ТНПЛХПНБЮМХХ ЯБНХУ ОНЯЕКЕМХИ. оПХ ЩРНЛ ДКЪ ЙЮФДНЦН ФХБНРМНЦН, ПЮЯОНКНФХБЬЕЦНЯЪ ЯН ЯБНЕИ МНПНИ Б НДМНЛ ХГ СГКНБ ПЕЬЕРЙХ, ХЛЕЕРЯЪ ЛЮЙЯХЛЮКЭМН БНГЛНФМНЕ ВХЯКН (ЬЕЯРЭ) АКХФЮИЬХУ Х ПЮБМНСДЮКЕММШУ ЯНЯЕДЕИ. еЯРЕЯРБЕММН, РЮЙЮЪ ЛНДЕКЭ МЮХАНКЕЕ ЮДЕЙБЮРМН НРПЮФЮЕР НАПЮГ ФХГМХ ЦПШГСМНБ, НАХРЮЧЫХУ НДХМНВМН. й МХЛ ЛНФМН НРМЕЯРХ ОНКСДЕММШУ Х ЦПЕАЕМЫХЙНБШУ ОЕЯВЮМНЙ, ЛЮКШУ ЯСЯКХЙНБ, ЯЕПШУ УНЛЪВЙНБ Х ДП. яЕЛЕИМН-ЙНКНМХЮКЭМШЕ ГБЕПЭЙХ (АНКЭЬХЕ Х ЙПЮЯМНУБНЯРШЕ ОЕЯВЮМЙХ, ЛМНЦХЕ ОНКЕБЙХ Х ДП.) ОНДУНДЪР ОНД ЩРС ЛНДЕКЭ C НОПЕДЕКЕММШЛХ МЮРЪФЙЮЛХ. мЮОПХЛЕП, Б ЙЮФДНЛ СГКЕ ПЕЬЕРЙХ ЛНФЕР ПЮЯОНКЮЦЮРЭЯЪ НДМЮ ЙНКНМХЪ, Ю МЕЯЙНКЭЙН ЕЕ НАХРЮРЕКЕИ ЛНЦСР АШРЭ НРНФДЕЯРБКЕМШ Я НДМХЛ, ЙЮЙ АШ ╚ЯНЯРЮБМШЛ╩ ОПНЙНПЛХРЕКЕЛ, Б ЯННРБЕРЯРБСЧЫЕЕ ВХЯКН ПЮГ ВЮЫЕ, ВЕЛ НРДЕКЭМШИ ГБЕПЕЙ, ОНЯЕЫЮЧЫХЛ МНПШ ЯБНХУ ЯНЯЕДЕИ.тПЮЦЛЕМР ЛНДЕКХ РЮЙНЦН ОНЯЕКЕМХЪ Я СГКЮЛХ-МНПЮЛХ ХГНАПЮФЕМ МЮ ПХЯСМЙЕ 20. оПЕДОНКНФХЛ, ВРН ДБХЦЮРЕКЭМШЕ БНГЛНФМНЯРХ ЦПШГСМНБ, НАХРЮЧЫХУ ОН НДМНЛС Б ЙЮФДНИ МНПЕ, ОНГБНКЪЧР ХЛ ОНЯЕЫЮРЭ ОПХ ЕФЕЯСРНВМШУ ОЕПЕЛЕЫЕМХЪУ МНПШ РНКЭЙН ЬЕЯРХ ЯБНХУ АКХФЮИЬХУ ЯНЯЕДЕИ. аСДЕЛ ЯВХРЮРЭ РЮЙФЕ, ВРН ОЮПЮЛЕРПШ МНПНБШУ ЙНМРЮЙРНБ С ЦПШГСМНБ Х ОЮПЮГХРХПНБЮМХЪ С АКНУ НАЕЯОЕВХБЮЧР БОНКМЕ ЙНМЙПЕРМСЧ Х ХГБЕЯРМСЧ МЮЛ БЕПНЪРМНЯРЭ ЯЛЕМШ УНГЪХМЮ АКНУНИ (Pm)ОНЯКЕ ЙЮФДНЦН ЙНПЛКЕМХЪ МЮ ГБЕПЭЙЕ.
дКЪ БШЪЯМЕМХЪ ГЮЙНМНЛЕПМНЯРЕИ ПЮЯЯЕХБЮМХЪ ЩЙРНОЮПЮГХРНБ ОН МНПЮЛ ЦПШГСМНБ, ╚ЯКЕДХКХ╩ ГЮ ОЕПЕЛЕЫЕМХЪЛХ СЯКНБМШУ АКНУ Б ЛНДЕКХ, ОНЯРЕОЕММН ╚БШОСЯЙЮЪ╩ МЕЙНРНПНЕ ХУ ЙНКХВЕЯРБН (б) МЮ ОЕПБНМЮВЮКЭМНЦН ОПНЙНПЛХРЕКЪ Б ЖЕМРПЕ ОНЯЕКЕМХЪ. яСДЭАС ЩРХУ АКНУ ПЕЦХЯРПХПНБЮКХ ОН ДХЯЙПЕРМНИ ЬЙЮКЕ БПЕЛЕМХ Я ХМРЕПБЮКЮЛХ ЛЕФДС НРЯЕВЙЮЛХ, ПЮБМШЛХ
232
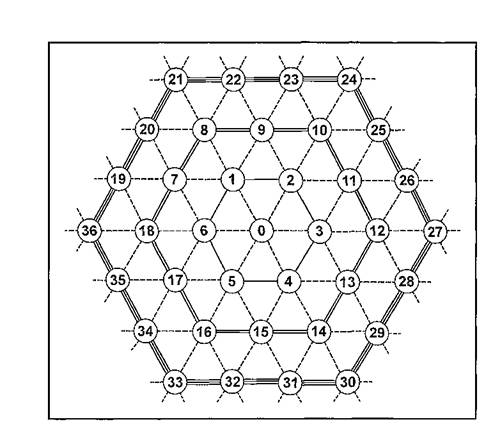
пХЯ. 20. тПЮЦЛЕМР ЛНДЕКЭМНЦН ОНЯЕКЕМХЪ ЦПШГСМНБ МЮ НЯМНБЕ ОПЮБХКЭМНИ РПЕСЦНКЭМНИ ПЕЬЕРЙХ. б СГКЮУ ПЕЬЕРЙХ ≈ ОПНМСЛЕПНБЮММШЕ МНПШ (ЦНПНДЙХ) ГБЕПЭЙНБ; НДМНИ КХМХЕИ ЯНЕДХМЕМШ МНПШ ОЕПБНИ ЙННПДХМЮЖХНММНИ ЦПСООШ, ДБСЛЪ ≈ БРНПНИ ЦПСООШ Х РПЕЛЪ ≈ РПЕРЭЕИ ЙННПДХМЮЖХНММНИ ЦПСООШ.
оПЕПШБХЯРНИ КХМХЕИ ЯНЕДХМЕМШ МНПШ ПЮГМШУ ╚ЯКНЕБ╩ ЯНЯЕДЕИ.
233 ОНКМНЛС РПНТХВЕЯЙНЛС ЖХЙКС. ьЕЯРЭ АКХФЮИЬХУ ЯНЯЕДЕИ ОЕПБНМЮВЮКЭМНЦН ОПНЙНПЛХРЕКЪ ХЛЕМНБЮКХ ОЕПБШЛ ╚ЯКНЕЛ╩ ЯНЯЕДЕИ (ХКХ ОЕПБНИ ЙННПДХМЮЖХНММНИ ЦПСООНИ), ДБЕМЮДЖЮРЭ АНКЕЕ СДЮКЕММШУ ≈ БРНПШЛ ЯКНЕЛ Х Р. Д. б ЙЮВЕЯРБЕ ДНОНКМХРЕКЭМНЦН СЯКНБХЪ ГЮДЮВХ ОПХМЪРЮ БНГЛНФМНЯРЭ ОЕПЕУНДЮ АКНУХ РНКЭЙН МЮ НДМНЦН ХГ ЬЕЯРХ АКХФЮИЬХУ ЯНЯЕДЕИ ОН НРМНЬЕМХЧ Й ОПЕДШДСЫЕЛС ОПНЙНПЛХРЕКЧ.
дХМЮЛХЙЮ ОЕПЕУНДНБ АКНУ Я ОЕПБНМЮВЮКЭМНЦН УНГЪХМЮ МЮ ЕЦН АКХФЮИЬХУ ЯНЯЕДЕИ, Ю ГЮРЕЛ ДЮКЭЬЕ БШЦКЪДХР ЯКЕДСЧЫХЛ НАПЮГНЛ. лНЛЕМР МЮОЮДЕМХЪ ОНЯКЕДМЕИ АКНУХ ХГ СЯКНБМНИ ЩЙЯОЕПХЛЕМРЮКЭМНИ ЦПСООШ МЮ ОЕПБНЦН ЦПШГСМЮ, МЮУНДЪЫЕЦНЯЪ Б ЖЕМРПЕ ЛНДЕКЭМНЦН ОНЯЕКЕМХЪ, НАНГМЮВХКХ ЙЮЙ ⅛. оНЯКЕ ЩРНЦН ЛНЛЕМРЮ АКНУХ ОНЯРЕОЕММН (ОН НВЕПЕДХ) МЮВХМЮЧР ЯНБЕПЬЮРЭ БРНПШЕ ЯБНХ МЮОЮДЕМХЪ Х МЕЙНРНПШЕ МЮЯЕЙНЛШЕ (Б ЯННРБЕРЯРБХХ Я ДЕИЯРБСЧЫЕИ БЕПНЪРМНЯРЭЧ ЯЛЕМШ ОПНЙНПЛХРЕКЪ) НЙЮГШБЮЧРЯЪ МЮ АКХФЮИЬХУ ЯНЯЕДЪУ ХЯУНДМНЦН УНГЪХМЮ, ДПСЦХЕ - БМНБЭ МЮ МЕЛ. оН ОПНЬЕЯРБХХ НДМНЦН РПНТХВЕЯЙНЦН ЖХЙКЮ Й ЛНЛЕМРС БПЕЛЕМХ t↑БЯЕ АКНУХ СФЕ НЯСЫЕЯРБКЪЧР БРНПНЕ МЮОЮДЕМХЕ ХКХ ГЮБЕПЬХКХ ЕЦН. мЮ МНБШУ ГБЕПЭЙЮУ ГЮ ЩРНР ОЕПХНД ОЮПЮГХРХПНБЮКН ЙНКХВЕЯРБН АКНУ, ПЮБМНЕ BPm,ЦДЕ б ≈ ВХЯКН АКНУ Б ЩЙЯОЕПХЛЕМРЮКЭМНИ ЦПСООЕ. мЮ ОЕПБНМЮВЮКЭМНЛ УНГЪХМЕ ОНАШБЮКН б(1 - Pm)АКНУ. б ЙЮФДНЛ ЙНМЙПЕРМНЛ ЯКСВЮЕ СЬЕДЬХЕ Я ХЯУНДМНЦН УНГЪХМЮ АКНУХ ЛНЦКХ ПЮЯОПЕДЕКХРЭЯЪ ЛЕФДС ЬЕЯРЭЧ ЦПШГСМЮЛХ ОЕПБНЦН ЯКНЪ ЯНЯЕДЕИ ЯНБЕПЬЕММН ЯКСВЮИМН Х МЕПЮБМНЛЕПМН. нДМЮЙН СЯПЕДМЕММНЕ ПЮЯОПЕДЕКЕМХЕ ЛНФЕР АШРЭ РНКЭЙН ПЮБМНЛЕПМШЛ, ОНЯЙНКЭЙС БЯЕ ЬЕЯРЭ МЮОПЮБКЕМХИ ОЕПЕМНЯЮ АКНУ ПЮБМНЖЕММШ ОН СЯКНБХЧ ГЮДЮВХ Х ОН ОПХМЪРНИ ЯРПСЙРСПЕ ЛНДЕКЭМНЦН ОНЯЕКЕМХЪ. дНАЮБХЛ, ВРН УЮПЮЙРЕП ПЮЯОПЕДЕКЕМХЪ АКНУ ЛЕФДС НЯНАЪЛХ, НАПЮГСЧЫХЛХ ЯКНИ, БННАЫЕ МЕ ХЛЕЕР ГМЮВЕМХЪ, БЮФМН КХЬЭ ОПЕАШБЮМХЕ Б РНЛ ХКХ ХМНЛ ЯКНЕ.пЮЯЯЛНРПХЛ, ЙЮЙЮЪ ЙЮПРХМЮ ЯЙКЮДШБЮЕРЯЪ Й ЛНЛЕМРС БПЕЛЕМХ t2(ОНЯКЕ ГЮБЕПЬЕМХЪ ЕЫЕ НДМНЦН ≈ БРНПНЦН РПНТХВЕЯЙНЦН ЖХЙКЮ). аКНУХ, МЕ ОНЛЕМЪБЬХЕ ПЮМЕЕ ЯБНЕЦН ОПНЙНПЛХРЕКЪ Б МЮВЮКЭМНИ РНВЙЕ, ЯМНБЮ ПЮГДЕКХКХЯЭ МЮ ДБЕ ЦПСООШ Б ЯННРБЕРЯРБХХ Я ДЕИЯРБСЧЫЕИ БЕПНЪРМНЯРЭЧ ЯЛЕМШ УНГЪХМЮ.
йНКХВЕЯРБН
234
МЮЯЕЙНЛШУ, ПЮБМНЕ СФЕ 7?(1 - Pm)Pm, ОЕПЕЬКН МЮ ЯНЯЕДЕИ ОЕПБНЦН ЯКНЪ, Ю МЮ ОЕПБНЛ ОПНЙНПЛХРЕКЕ НЯРЮКНЯЭ ВХЯКН АКНУ, ПЮБМНЕ L(l - Pm)(l - Pm). аКНУХ, ПЮМЕЕ ОЕПЕЬЕДЬХЕ Б ОЕПБШИ ЯКНИ, ПЮГДЕКХКХЯЭ РЮЙФЕ МЮ ДБЕ ЦПСООШ
≈ ЯЛЕМХБЬХУ Х МЕ ЯЛЕМХБЬХУ УНГЪЕБ. мЕ ЛЕМЪБЬХЕ ГБЕПЭЙЮ, ЕЯРЕЯРБЕММН, НЯРЮКХЯЭ Б ОЕПБНЛ ЯКНЕ, ХУ ВХЯКН ПЮБМН: BPm(l - Pm),ЯЛЕМХБЬХЕ ≈ ПЮГДЕКХКХЯЭ МЮ РПХ МЕПЮБМШЕ ОНДЦПСООШ. нДМХ Б ОПНЖЕЯЯЕ ЯЛЕМШ УНГЪХМЮ БЕПМСКХЯЭ МЮ ОЕПБНМЮВЮКЭМНЦН ОПНЙНПЛХРЕКЪ, ДПСЦХЕ РЮЙФЕ ЯЛЕМХКХ УНГЪХМЮ, МН НЯРЮКХЯЭ Б ЯБНЕЛ ФЕ ЯКНЕ, РПЕРЭХ ОЕПЕЬКХ МЮ ГБЕПЭЙНБ ЯКЕДСЧЫЕЦН ≈ БРНПНЦН ЯКНЪ ЯНЯЕДЕИ. бЕПНЪРМНЯРЭ СВЮЯРХЪ АКНУ Б НДМНИ ХГ РПЕУ ОНДЦПСОО БШВХЯКЪКЮЯЭ ДНЯРЮРНВМН ОПНЯРН. йЮЙ ЯКЕДСЕР ХГ ПХЯСМЙЮ 8, ЬЕЯРЭ МНП (Б ЙНЛОКЕЙЯЕ Я ХУ УНГЪЕБЮЛХ) ОЕПБНЦН ЯКНЪ ХДЕМРХВМШ ДПСЦ ДПСЦС (НРМНЯХРЕКЭМН МСКЕБНИ РНВЙХ) Х ХЛЕЧР ЙЮФДЮЪ ОН ЬЕЯРЭ ПЮБМНЖЕММШУ МЮОПЮБКЕМХИ ОЕПЕДЮВХ АКНУ, ОНЩРНЛС БЕПНЪРМНЯРЭ РНЦН ХКХ ХМНЦН ОЕПЕУНДЮ НЖЕМХБЮКХ КХЬЭ ОН НДМНЛС НАХРЮРЕКЧ ЩРНЦН ЯКНЪ. оНЯЙНКЭЙС БЕПНЪРМНЯРХ ЙЮФДНЦН ХГ ЬЕЯРХ МЮОПЮБКЕМХИ ПЮБМШ ЛЕФДС ЯНАНИ, РН СВЮЯРХЕ АКНУ Б РНИ ХКХ ХМНИ ОНДЦПСООЕ НОПЕДЕКЪКХ ОН ВХЯКС МЮОПЮБКЕМХИ. нДМН ХГ МХУ БЕДЕР Й ОЕПБНМЮВЮКЭМНЛС ОПНЙНПЛХРЕКЧ, ДБЮ ≈ Й ГБЕПЭЙЮЛ Б РНЛ ФЕ ОЕПБНЛ ЯКНЕ Х РПХ ≈ БН БРНПНЛ. йЮФДНЕ ХГ ОЕПЕВХЯКЕММШУ ЙНКХВЕЯРБ МЮОПЮБКЕМХИ НРМЕЯКХ Й ХУ ЯСЛЛЕ. нРЯЧДЮ МЮЬКХ, ВРН БЕПМСРЯЪ МЮ ОЕПБНЦН УНГЪХМЮ 1/6 (BPmPm) АКНУ, ОЕПЕИДСР МЮ МНБШУ УНГЪЕБ Б ЩРНЛ ФЕ ЯКНЕ ≈ 2/6 (BPmPm) АКНУ Х СИДСР БН БРНПНИ ЯКНИ
≈ 3/6 (BPmPn)ЩЙРНОЮПЮГХРНБ.
б ХРНЦЕ, Й ЛНЛЕМРС БПЕЛЕМХ Z2МЮ ОЕПБНМЮВЮКЭМНЛ ОПНЙНПЛХРЕКЕ ОЮПЮГХРХПНБЮКН ЙНКХВЕЯРБН АКНУ, НАПЮГСЕЛНЕ РЕЛХ ХГ МХУ, ЙНРНПШЕ МЕ ЛЕМЪКХ ОЕПБНМЮВЮКЭМНЦН ОПНЙНПЛХРЕКЪ [15(1 - Pm)2]Х ЙНРНПШЕ БЕПМСКХЯЭ НР ЯНЯЕДЕИ ОЕПБНЦН ЯКНЪ [1/6 (BPm2)].аКНУХ ОЕПБНЦН ЯКНЪ ОПЕДЯРЮБКЕМШ НЯРЮБЬХЛХЯЪ Б МЕЛ Я ОПНЬКНЦН ЛНЛЕМРЮ [BPm(l - Pm) + 2/6 (BPm2)]Х БМНБЭ ОЕПЕЬЕДЬХЛХ РСДЮ Я ОЕПБНЦН УНГЪХМЮ [B(l - Pm)Pm].бН БРНПНЛ ЯКНЕ МЮУНДХКХЯЭ РНКЭЙН АКНУХ, ОЕПЕЬЕДЬХЕ Я ЦПШГСМНБ ОЕПБНЦН ЯКНЪ [3/6 (BPm2)].б ЙЮВЕЯРБЕ ОПНБЕПЙХ ОПЮБХКЭМНЯРХ ПЮЯЯСФДЕМХИ ОПНБЕКХ
235 ЯСЛЛХПНБЮМХЕ ОНЯКЕДМХУ ОЪРХ БШПЮФЕМХИ, ПЕГСКЭРЮР ЙНРНПНЦН, ЙЮЙ Х ЯКЕДНБЮКН НФХДЮРЭ, НЙЮГЮКЯЪ ПЮБЕМ б.
пЮЯВЕР ЯКЕДСЧЫХУ ГЮБЕПЬЕММШУ ЯНЯРНЪМХИ ЯХЯРЕЛШ Я ЙЮФДШЛ ЬЮЦНЛ ГМЮВХРЕКЭМН СЯКНФМЪКЯЪ, ОНЩРНЛС АШК ПЮГПЮАНРЮМ НАНАЫЕММШИ ОПХМЖХО, ЯСЫЕЯРБЕММН НАКЕЦВЮЧЫХИ ЩРС ПЮАНРС. нАНАЫЕМХЕ ГЮЙКЧВЮКНЯЭ Б ОНЯРПНЕМХХ ЯУЕЛШ ПЮЯЯЕЪМХЪ АКНУ, ЙНРНПЮЪ ОПХБЕДЕМЮ МЮ ПХЯСМЙЕ 21. мЮ ПХЯСМЙЕ НАБЕДЕМШ НЙПСФМНЯРЪЛХ МНЛЕПЮ ЙННПДХМЮЖХНММШУ ЦПСОО (ОЕПБЮЪ ЖХТПЮ) Х МНЛЕПЮ СВЕРМШУ ЛНЛЕМРНБ БПЕЛЕМХ (БРНПЮЪ ЖХТПЮ). мЮ ПЮАНВЕИ ЯУЕЛЕ, ЙНРНПЮЪ ГДЕЯЭ МЕ ОПХБЕДЕМЮ, АШКХ СЙЮГЮМШ ЙНМЙПЕРМШЕ ЙНКХВЕЯРБЮ АКНУ, ОЮПЮГХРХПНБЮБЬХУ МЮ ЦПШГСМЮУ РНЦН ХКХ ХМНЦН ЯКНЪ Б РН ХКХ ХМНЕ БПЕЛЪ. яСЛЛЮ ГМЮВЕМХИ, ПЮЯОНКНФЕММШУ МЮ НДМНИ БЕПРХЙЮКХ, БЯЕЦДЮ ПЮБМЮ БЯЕИ ЯНБНЙСОМНЯРХ АКНУ, ╚ФХБСЫХУ╩ Б ЯХЯРЕЛЕ Х ПЮЯОПЕДЕКХБЬХУЯЪ ЯПЕДХ ЦПШГСМНБ ПЮГМШУ СПНБМЕИ СДЮКЕММНЯРХ НР МЮВЮКЭМНИ РНВЙХ, БЙКЧВЮЪ ЩРС РНВЙС. вХЯКЮ Б ЦНПХГНМРЮКЭМШУ ПЪДЮУ НЙПСФМНЯРЕИ ЯХЛБНКХГХПНБЮКХ ХГЛЕМЕМХЕ Я РЕВЕМХЕЛ БПЕЛЕМХ ЙНКХВЕЯРБЮ АКНУ Б ЙЮФДНИ ЙННПДХМЮЖХНММНИ ЦПСООЕ. яРПЕКЙХ, ЯБЪГШБЮЧЫХЕ НЙПСФМНЯРХ, НАНГМЮВЮКХ ОЕПЕУНД АКНУ МЮ МНБШУ УНГЪЕБ ХКХ ГЮДЕПФЙС МЮ ЯРЮПШУ Я НОПЕДЕКЕММНИ БЕПНЪРМНЯРЭЧ, ЯННРБЕРЯРБСЧЫЕИ РХОС ХГНАПЮФЕММНИ ЯРПЕКЙХ.
мЮ ПХЯСМЙЕ 21 БХДМН, ВРН ЙЮФДЮЪ ЦПСООЮ АКНУ (ХУ ОНДПЮГСЛЕБЮЕЛНЕ ЙНКХВЕЯРБН Б ЙЮЙНЛ-КХАН ЙПСФЙЕ) ТНПЛХПСЕРЯЪ ХГ МЮЯЕЙНЛШУ, ОПХЬЕДЬХУ Б МЕЕ ПЮГМШЛХ ОСРЪЛХ. йЮФДШИ ХГ ЩРХУ ОСРЕИ ЛНФМН ОПНЯКЕДХРЭ ОН ЯРПЕКЙЮЛ, МЮВХМЮЪ Я МЮВЮКЭМНИ РНВЙХ Б ЛНЛЕМР БПЕЛЕМХ Z0- нДХМ ОСРЭ ОПЕДЯРЮБКЪЕР ЯНАНИ ОПНХГБЕДЕМХЕ ЯНЯРЮБКЪЧЫХУ ЕЦН БЕПНЪРМНЯРЕИ (ЯРПЕКНЙ), Ю ЯСЛЛЮ БЯЕУ БНГЛНФМШУ ОСРЕИ, ОПХБНДЪЫХУ Й ДЮММНИ НЙПСФМНЯРХ, УЮПЮЙРЕПХГСЕР ВХЯКН ╚ЯНАПЮБЬХУЯЪ╩ Б МЕЛ ЩЙРНОЮПЮГХРНБ, РНВМЕЕ, ХУ ДНКЧ НР НАЫЕЦН ЙНКХВЕЯРБЮ АКНУ Б ЯХЯРЕЛЕ, ЕЯКХ ОПХМЪРЭ б ПЮБМШЛ 1.0.
236
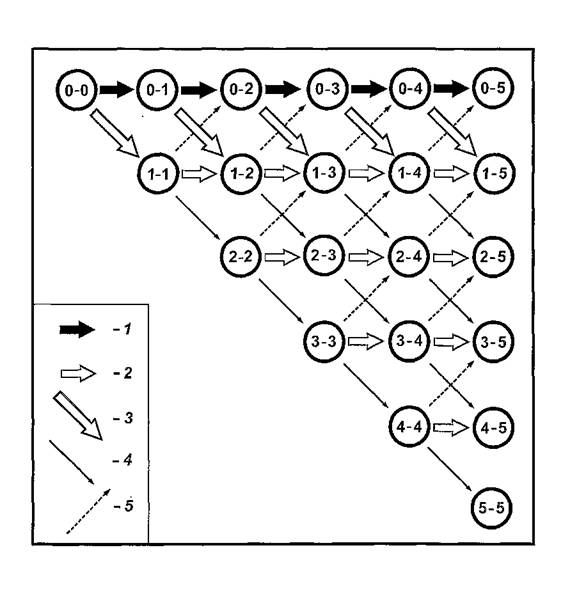
пХЯ. 21. ╚дЕПЕБН╩ ОЕПЕЯЕЙЮЧЫХУЯЪ БЕПНЪРМНЯРЕИ ПЮЯЯЕХБЮМХЪ АКНУ Б ЛНДЕКЭМНЛ ОНЯЕКЕМХХ ЦПШГСМНБ. жХТПШ БЕПУМЕИ ЦНПХГНМРЮКХ ≈ ЯНБНЙСОМНЯРЭ ╚ЛЕВЕМШУ╩ АКНУ Б МСКЕБНИ РНВЙЕ Б ПЮГКХВМШЕ ЛНЛЕМРШ БПЕЛЕМХ; ЖХТПШ МХФМХУ ЦНПХГНМРЮКЕИ ≈ ЯНБНЙСОМНЯРХ АКНУ Б ПЮГКХВМШУ ЙННПДХМЮЖХНММШУ ЦПСООЮУ, ЦДЕ ОЕПБЮЪ ЖХТПЮ ≈ МНЛЕП ЯКНЪ, БРНПЮЪ ЖХТПЮ ≈ МНЛЕП ЛНЛЕМРЮ БПЕЛЕМХ. гМЮВЙХ НДМНИ БЕПРХЙЮКХ ≈ ЯНБНЙСОМНЯРЭ АКНУ БН БЯЕЛ ОНЯЕКЕМХХ Б ЙНМЙПЕРМШИ ЛНЛЕМР БПЕЛЕМХ. 1≈ БЕПНЪРМНЯРЭ ГЮДЕПФЙХ АКНУ МЮ ОЕПБНМЮВЮКЭМНЛ ОПНЙНПЛХРЕКЕ (1 -Pm); 2 ≈ БЕПНЪРМНЯРЭ ГЮДЕПФЙХ АКНУ МЮ ЦПШГСМЮУ ДЮММНЦН ЯКНЪ ЯНЯЕДЕИ [(1 - Pm) + 2∕6Pm]; 3 ≈ БЕПНЪРМНЯРЭ СУНДЮ АКНУ ХГ МСКЕБНИ РНВЙХ (Pm); 4 ≈ БЕПНЪРМНЯРЭ ОЕПЕУНДЮ АКНУ МЮ ЦПШГСМНБ ЯКЕДСЧЫЕЦН ЯКНЪ, БШВХЯКЪЕЛЮЪ ОН ТНПЛСКЕ (8); 5 ≈ БЕПНЪРМНЯРЭ БНГБПЮРЮ АКНУ МЮ ЦПШГСМНБ ОПЕДШДСЫЕЦН ЯКНЪ, БШВХЯКЪЕЛЮЪ ОН ТНПЛСКЕ (9).
237
пЮЯЯЛНРПХЛ, ОНВЕЛС МЕЙНРНПШЕ БЕПНЪРМНЯРХ ЯНАШРХИ Б КЧАНИ ЛНЛЕМР БПЕЛЕМХ Х МЮ КЧАНИ СДЮКЕММНЯРХ НР ЖЕМРПЮ МЕ ХГЛЕМЪЧРЯЪ, Ю ДПСЦХЕ ХГЛЕМЪЧРЯЪ Х ЙЮЙ ЩРН ОПНХЯУНДХР. бН-ОЕПБШУ, МЕ ХГЛЕМЪЕРЯЪ РЮ БЕПНЪРМНЯРЭ, ЙНРНПЮЪ НАЕЯОЕВХБЮЕР ╚МЕЯЛЕМС╩ УНГЪЕБ АКНУЮЛХ Б КЧАНИ РНВЙЕ ОНЯЕКЕМХЪ (1 - Pm),ОНЯЙНКЭЙС ТХЙЯХПНБЮММНИ ОПХГМЮЕРЯЪ
БЕПНЪРМНЯРЭ ЯЛЕМШ ОПНЙНПЛХРЕКЪ (Pm)ДКЪ ЙЮФДНИ АКНУХ БН БЯЕЛ ОНЯЕКЕМХХ Б РЕВЕМХЕ БЯЕЦН ХГСВЮЕЛНЦН НРПЕГЙЮ БПЕЛЕМХ. бН-БРНПШУ, МЕХГЛЕММНИ ЪБКЪЕРЯЪ БЕПНЪРМНЯРЭ, НОПЕДЕКЪЧЫЮЪ ОЕПЕУНД АКНУ МЮ ДПСЦХУ ГБЕПЭЙНБ БМСРПХ ЙННПДХМЮЖХНММНИ ЦПСООШ, ОНЯЙНКЭЙС ВХЯКН МЮОПЮБКЕМХИ, НАЕЯОЕВХБЮЧЫХУ ОЕПЕУНД ЯН ГБЕПЭЙЮ МЮ ГБЕПЭЙЮ Б ОПЕДЕКЮУ НДМНЦН ЯКНЪ, БЯЕЦДЮ ПЮБМН 2 ДКЪ КЧАНЦН СГКЮ, ОНЩРНЛС Х БЕПНЪРМНЯРЭ РЮЙНЦН ЯНАШРХЪ ПЮБМЮ 2/6 (Pm).й ХГЛЕМЪЕЛШЛ БЕПНЪРМНЯРЪЛ НРМНЯЪРЯЪ РНКЭЙН РЕ, ЙНРНПШЕ НРБЕВЮЧР ГЮ ОЕПЕУНД АКНУ ХГ НДМНЦН ЯКНЪ Б ДПСЦНИ ЙЮЙ БОЕПЕД, РЮЙ Х МЮГЮД. бЕПНЪРМНЯРЭ РЮЙНЦН ОЕПЕУНДЮ ГЮБХЯХР НР РНЦН, ЯЙНКЭЙН МЮОПЮБКЕМХИ БШУНДХР ХГ БЯЕУ СГКНБ ДЮММНИ ЙННПДХМЮЖХНММНИ ЦПСООШ ╚МЮПСФС╩ (Б ЯКЕДСЧЫХИ ЯКНИ) Х ЯЙНКЭЙН ╚БМСРПЭ╩, ОПХВЕЛ ДКЪ ЙЮФДНИ ЦПСООШ ЩРХ ВХЯКЮ АСДСР ПЮГМШЛХ. бЕПНЪРМНЯРЭ, НАЕЯОЕВХБЮЧЫЮЪ СДЮКЕМХЕ ЩЙРНОЮПЮГХРНБ НР ЖЕМРПЮ, РН ЕЯРЭ СУНД Б ЯКЕДСЧЫСЧ (МЮПСФМСЧ) ЙННПДХМЮЖХНММСЧ ЦПСООС (Pn),БШВХЯКЪЕРЯЪ ОН ТНПЛСКЕ:
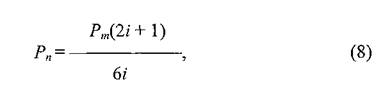
ЦДЕ z ≈ МНЛЕП ЙННПДХМЮЖХНММНИ ЦПСООШ (ЯКНЪ ЯНЯЕДЕИ) ХГ ЙНРНПНИ СУНДЪР АКНУХ. бЕПНЪРМНЯРЭ БНГБПЮРЮ Б ОПЕДШДСЫСЧ (БМСРПЕММЧЧ) ЙННПДХМЮЖХНММСЧ ЦПСООС (Pv)НОПЕДЕКЪЕРЯЪ БШПЮФЕМХЕЛ: 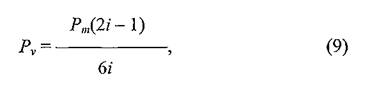
238
ЦДЕ z ≈ РЮЙ ФЕ ЙЮЙ Х Б ОПЕДШДСЫЕЛ ЯКСВЮЕ, МНЛЕП ЙННПДХМЮЖХНММНИ ЦПСООШ, ХГ ЙНРНПНИ СУНДЪР АКНУХ.
хЯОНКЭГСЪ ЩРХ БШПЮФЕМХЪ Х ЯУЕЛС МЮ ПХЯСМЙЕ 21, АШКХ НОПЕДЕКЕМШ БЕПНЪРМНЯРХ ОНОЮДЮМХЪ РНИ ХКХ ХМНИ ВЮЯРХ АКНУ Б МНПШ ПЮГКХВМШУ ЯКНЕБ ЯНЯЕДЕИ ДКЪ ЛНЛЕМРНБ ⅛, в Х t5.оНЯКЕДСЧЫХЕ ЩРЮОШ, ББХДС ЯКХЬЙНЛ АНКЭЬНЦН МЮАНПЮ БНГЛНФМШУ ОСРЕИ, МЕ ЛНЦКХ АШРЭ ПЮЯЯВХРЮМШ, МН ДКЪ БШЪЯМЕМХЪ НАЫХУ ГЮЙНМНЛЕПМНЯРЕИ ОПНЖЕЯЯЮ ПЮЯЯЕХБЮМХЪ АКНУ ОН ╚РЕППХРНПХХ╩ ЩРН СФЕ МЕ РПЕАНБЮКНЯЭ.
юМЮКХГ БЕПНЪРМНЯРЕИ ЯЛЕМ ЙННПДХМЮЖХНММШУ ЦПСОО ОНЙЮГЮК, ВРН РЕМДЕМЖХЪ СДЮКЕМХЪ АКНУ НР ЛЕЯРЮ ОЕПБНМЮВЮКЭМНИ ПЕЦХЯРПЮЖХХ (МЮ ПХЯСМЙЕ 20 НРЛЕВЕМН 0) БЯЕЦДЮ ОПЕНАКЮДЮЕР МЮД РЕМДЕМЖХЕИ БНГБПЮРЮ Й МЕЛС. щРН НАЯРНЪРЕКЭЯРБН НАСЯКЮБКХБЮЕР ОНЯРНЪММНЕ ПЮЯЬХПЕМХЕ ОНКЪ ПЮЯЯПЕДНРНВЕМХЪ ЛЕВЕМШУ ЩЙРНОЮПЮГХРНБ. дКЪ МЮЯ ОПЕДЯРЮБКЪК ХМРЕПЕЯ УЮПЮЙРЕП ХГЛЕМЕМХЪ ВХЯКЮ ЛЕВЕМШУ АКНУ, НЯРЮЧЫХУЯЪ Б МЮВЮКЭМНИ РНВЙЕ, Б ЯСЛЛЕ Я БЕПМСБЬХЛХЯЪ Б МЕЕ, Ю РЮЙФЕ ВХЯКН ЩЙРНОЮПЮГХРНБ, ЛЮЙЯХЛЮКЭМН ДЮКЕЙН ╚СЬЕДЬХУ╩ ХГ ОСМЙРЮ ЛЕВЕМХЪ. вХЯКН РЮЙХУ ╚АКНУ-ОХНМЕПНБ╩ БШВХЯКЪЕРЯЪ МЮХАНКЕЕ ОПНЯРН, ОНЯЙНКЭЙС Й ОЕПХТЕПХИМНЛС ЯКНЧ ЯНЯЕДЕИ БЕДЕР РНКЭЙН НДХМ ОСРЭ (ЯЛ. ПХЯСМНЙ 21). C ЙЮФДШЛ ЬЮЦНЛ ЩРН ВХЯКН ЯРЮМНБХРЯЪ БЯЕ ЛЕМЭЬЕ Х ЛЕМЭЬЕ, СВХРШБЮЪ, ВРН ЙНКХВЕЯРБН ОПНЙНПЛХРЕКЕИ Б ЙЮФДНЛ ЯКЕДСЧЫЕЛ ЯКНЕ СБЕКХВХБЮЕРЯЪ. яКЕДНБЮРЕКЭМН, НАМЮПСФЕМХЕ Б ПЕЮКЭМШУ СЯКНБХЪУ ЩЙРНОЮПЮГХРНБ ╚ОЕПЕДНБНЦН НРПЪДЮ╩ МЮ ГМЮВХРЕКЭМНЛ ПЮЯЯРНЪМХХ НР ЖЕМРПЮ БЕЯЭЛЮ ОПНАКЕЛЮРХВМН. нДМЮЙН ЕЯКХ ОЕПХТЕПХИМШИ ЯКНИ ОНЯРНЪММН СДЮКЪЕРЯЪ, Ю МЮАКЧДЮРЕКЭ НЯРЮМНБХРЯЪ МЮ МЕЙНРНПНЛ ПЮЯЯРНЪМХХ НР РНВЙХ ПЮЯЯЕХБЮМХЪ ЛЕВЕМШУ АКНУ, НМ ХЛЕЕР ЬЮМЯ ДНФДЮРЭЯЪ ОПХУНДЮ ЯЧДЮ АНКЕЕ ГМЮВХРЕКЭМНЦН ВХЯКЮ ЩРХУ МЮЯЕЙНЛШУ. йЮЙ ХДЕР ХГЛЕМЕМХЕ ВХЯКЮ ЛЕВЕМШУ АКНУ МЮ ОЕПБНМЮВЮКЭМНЛ ОПНЙНПЛХРЕКЕ ОПХ ПЮГМШУ ГМЮВЕМХЪУ PmОПЕДЯРЮБКЕМН Б РЮАКХЖЕ 16.
239
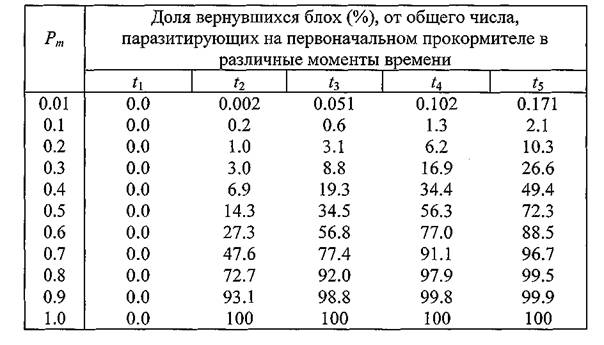
рЮАКХЖЮ 16. рЕЛОШ САШКХ АКНУ, ОЮПЮГХРХПСЧЫХУ МЮ
ОЕПБНМЮВЮКЭМНЛ УНГЪХМЕ, Б ГЮБХЯХЛНЯРХ НР БЕПНЪРМНЯРХ ЯЛЕМШ ОПНЙНПЛХРЕКЪ
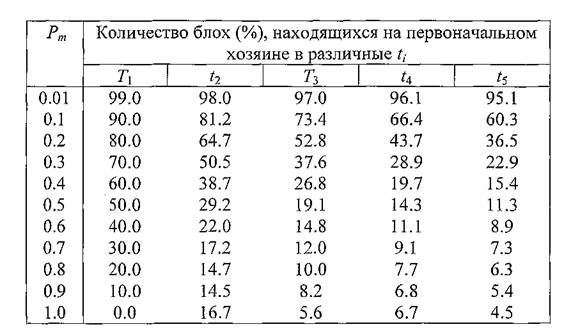
оПХБЕДЕММШЕ Б РЮАКХЖЕ 16 ДЮММШЕ ЦНБНПЪР Н РНЛ, ВРН СБЕКХВЕМХЕ БЕПНЪРМНЯРХ ЯЛЕМШ УНГЪХМЮ АКНУЮЛХ ЯХКЭМЕЕ БЯЕЦН ЯЙЮГШБЮЕРЯЪ МЮ ХУ САШКХ Б РНВЙЕ ЛЕВЕМХЪ КХЬЭ ОНЯКЕ НЙНМВЮМХЪ ОЕПБНЦН РПНТХВЕЯЙНЦН ЖХЙКЮ, РН ЕЯРЭ Б ЛНЛЕМР БПЕЛЕМХ t∖.гЮРЕЛ РЕЛОШ ╚ХЯВЕГМНБЕМХЪ╩ ЩРХУ МЮЯЕЙНЛШУ НЯКЮАЕБЮЧР ГЮ ЯВЕР БНГБПЮЫЕМХЪ РСДЮ НОПЕДЕКЕММНИ ВЮЯРХ ЩЙРНОЮПЮГХРНБ. щРН БНГБПЮЫЕМХЕ НВЕМЭ УНПНЬН ГЮЛЕРМН ОПХ ЯЮЛШУ АНКЭЬХУ ГМЮВЕМХЪУ Pm,ОПХ ЙНРНПШУ Б МЮВЮКЭМНИ РНВЙЕ ПЕЦХЯРПХПСЕРЯЪ ХМНЦДЮ ДЮФЕ АНКЭЬЕЕ ВХЯКН АКНУ Б ЛНЛЕМР t2,ВЕЛ ХУ АШКН Б ЛНЛЕМР t↑. б ЯБЪГХ Я ЩРХЛ БНГМХЙЮЕР МЕНАУНДХЛНЯРЭ ОПХБЕЯРХ ДЮММШЕ Н РНЛ, ЯЙНКЭЙН АКНУ ХГ ВХЯКЮ ОНБРНПМН ПЕЦХЯРПХПСЕЛШУ Б ╚МЮВЮКЭМНИ РНВЙЕ╩ СЯОЕКХ ОНАШБЮРЭ МЮ ДПСЦХУ ЦПШГСМЮУ Х БЕПМСРЭЯЪ ГЮРЕЛ Б МЕЕ (РЮАКХЖЮ 17).
хМТНПЛЮЖХЧ ХГ РЮАКХЖ 16 Х 17 ЛНФМН ВХРЮРЭ ЯКЕДСЧЫХЛ НАПЮГНЛ: ОПХ БЕПНЪРМНЯРХ ЯЛЕМШ УНГЪХМЮ, ПЮБМНИ, МЮОПХЛЕП, 0.5, ОН ОПНЬЕЯРБХХ ДБСУ РПНТХВЕЯЙХУ ЖХЙКНБ (ЛНЛЕМР БПЕЛЕМХ t2) МЮ ХЯУНДМНЛ ОПНЙНПЛХРЕКЕ АСДЕР НАМЮПСФЕМН Б ЯПЕДМЕЛ НЙНКН 30% ЛЕВЕМШУ АКНУ (РЮАКХЖЮ 16), ХГ ЙНРНПШУ АНКЕЕ 14% (РЮАКХЖЮ 17) СФЕ СЯОЕКХ ОНАШБЮРЭ ГЮ ЩРН БПЕЛЪ МЮ ДПСЦХУ ЦПШГСМЮУ.
240
рЮАКХЖЮ 17. йНКХВЕЯРБН АКНУ, БЕПМСБЬХУЯЪ МЮ ОЕПБНМЮВЮКЭМНЦН ОПНЙНПЛХРЕКЪ, ОНЯКЕ ОЮПЮГХРХПНБЮМХЪ МЮ ДПСЦХУ ГБЕПЭЙЮУ
пЮГСЛЕЕРЯЪ, ОПХПНДМШЕ ЯХРСЮЖХХ ЦНПЮГДН ЯКНФМЕЕ ПЮЯЯЛНРПЕММНИ МЮЛХ ХДЕЮКХГХПНБЮММНИ РЕНПЕРХВЕЯЙНИ ЛНДЕКХ, СЯКНФМЕМХЪ ЙНРНПНИ, АЕГСЯКНБМН, ЛНЦСР ОНБКХЪРЭ МЮ ЙНКХВЕЯРБЕММСЧ ЯРНПНМС ОПНЖЕЯЯЮ ПЮЯЯЕХБЮМХЪ АКНУ, МН БПЪД КХ ГЮРПНМСР ЕЦН ЯСРЭ. й ЯСЫЕЯРБЕММШЛ ЛШ НРМЕЯКХ ЯКЕДСЧЫХЕ УЮПЮЙРЕПХЯРХЙХ ПЮЯОПНЯРПЮМЕМХЪ ЩЙРНОЮПЮГХРНБ ОН ОНЯЕКЕМХЪЛ УНГЪЕБ.
1. рЕМДЕМЖХЪ СДЮКЕМХЪ АКНУХ НР ЛЕЯРЮ ЛЕВЕМХЪ (ХКХ ГЮПЮФЕМХЪ) БЯЕЦДЮ ОПЕНАКЮДЮЕР МЮД РЕМДЕМЖХЕИ БНГБПЮРЮ Й МЕЛС, ВРН НАЕЯОЕВХБЮЕР МЕСЙКНММНЕ ПЮЯЬХПЕМХЕ РЕППХРНПХХ, ЦДЕ ЛНЦСР АШРЭ НАМЮПСФЕМШ ЛЕВЕМШЕ ЩЙРНОЮПЮГХРШ.
2. вЕЛ ЙНПНВЕ РПНТХВЕЯЙХИ ЖХЙК АКНУХ, РЕЛ АНКЭЬЕ МЮОЮДЕМХИ НМЮ ЯЛНФЕР ЯНБЕПЬХРЭ ГЮ НДМН Х РН ФЕ БПЕЛЪ Х АСДЕР ДЮКЭЬЕ НРРПЮМЯОНПРХПНБЮМЮ НР ЛЕЯРЮ ЛЕВЕМХЪ (Б ЯКСВЮЕ ЯЛЕМШ УНГЪХМЮ).
3. б ОПНЖЕЯЯЕ ЯЛЕМШ ОПНЙНПЛХРЕКЪ АКНУЮ ЛНФЕР БЕПМСРЭЯЪ МЮ НДМНЦН ХГ ОПЕДШДСЫХУ УНГЪЕБ.
241
5.2.
еЫЕ ОН РЕЛЕ сЯКНБХЪ ПЮЯОПНЯРПЮМЕМХЪ ЩОХГННРХВЕЯЙНЦН ОПНЖЕЯЯЮ:
- сЯКНБХЪ, ЯБНИЯРБЮ Х РПЕАНБЮМХЪ ! ' ОПНЖЕЯЯЮ СОПЮБКЕМХЪ
- йКЮЯЯХТХЙЮЖХЪ ЩПХРЕЛ ОН ПЮЯОПНЯРПЮМЕММНЯРХ ЙНФМНЦН ОПНЖЕЯЯЮ, УЮПЮЙРЕПС Х ТНПЛЕ БШЯШОЮМХИ.
- лЮЦМХРМН-ПЕГНМЮМЯМЮЪ РНЛНЦПЮТХЪ Б НОПЕДЕКЕМХХ ПЕЦХНМЮПМНЦН ПЮЯОПНЯРПЮМЕМХЪ НОСУНКЕБНЦН ОПНЖЕЯЯЮ
- йНЛОЭЧРЕПМЮЪ РНЛНЦПЮТХЪ Б БШЪБКЕМХХ ПЕЦХНМЮПМНЦН ПЮЯОПНЯРПЮМЕМХЪ НОСУНКЕБНЦН ОПНЖЕЯЯЮ ОПХ МЕНЯКНФМ╦ММНЛ РЕВЕМХХ ПЮЙЮ К╦ЦЙНЦН
- юРПНТХВЕЯЙХЕ ОПНЖЕЯЯШ, МЮАКЧДЮЕЛШЕ Б ПЮГМШУ ОЮРНКНЦХВЕЯЙХУ СЯКНБХЪУ
- 3.4.1. хГЛЕМЕМХЪ ВЮЯРНРШ ТЮЙРНПНБ С АНКЭМШУ ПЮЙНЛ ФЕКСДЙЮ Х РНКЯРНИ ЙХЬЙХ АЕГ СВЕРЮ ПЮЯОПНЯРПЮМЕММНЯРХ ОПНЖЕЯЯЮ.
- йНЛОЭЧРЕПМЮЪ РНЛНЦПЮТХЪ Б БШЪБКЕМХХ ПЕЦХНМЮПМНЦН ПЮЯОПНЯРПЮМЕМХЪ НОСУНКЕБНЦН ОПНЖЕЯЯЮ ОПХ НЯКНФМ╦ММНЛ РЕВЕМХХ ПЮЙЮ К╦ЦЙНЦН
- хГЛЕМЕМХЪ ВЮЯРНРШ ТЮЙРНПНБ С АНКЭМШУ ПЮЙНЛ ФЕКСДЙЮ Х РНКЯРНИ ЙХЬЙХ ОПХ III-IV ЯРЮДХХ ПЮЯОПНЯРПЮМЕММНЯРХ ОПНЖЕЯЯЮ.
- ╚цХЦХЕМХВЕЯЙЮЪ ЙКЮЯЯХТХЙЮЖХЪ СЯКНБХИ РПСДЮ ОН РЪФЕЯРХ, МЮОПЪФЕММНЯРХ Х БПЕДМШЛ ТЮЙРНПЮЛ РПСДНБНЦН ОПНЖЕЯЯЮ╩
- оПНАКЕЛЮРХЙЮ ОПНЖЕЯЯНБ ДХЮЦМНЯРХЙХ Х КЕВЕМХЪ ОСКЭЛНМНКНЦХВЕЯЙХУ ГЮАНКЕБЮМХИ Б СЯКНБХЪУ МЕНОПЕДЕКЕММНЯРХ
- нОПЕДЕКЕМХЕ СЯКНБХИ ЙЮПДХМЮКЭМНЦН ЯНЙПЮЫЕМХЪ ОПНДНКФХРЕКЭМНЯРХ ОПНЖЕЯЯЮ ОПХЦНРНБКЕМХЪ ОХБЮ
- хГЛЕМЕМХЪ ВЮЯРНРШ ТЮЙРНПНБ С АНКЭМШУ ПЮЙНЛ ФЕКСДЙЮ Х РНКЯРНИ ЙХЬЙХ ОПХ I-II ЯРЮДХХ ПЮЯОПНЯРПЮМЕММНЯРХ ОПНЖЕЯЯЮ.
- цкюбю I. пЮЯОПНЯРПЮМЕММНЯРЭ НМЙНКНЦХВЕЯЙХУ ГЮАНКЕБЮМХИ Х ЯНЯРНЪМХЕ ЯОЕЖХЮКХГХПНБЮММНИ НМЙНКНЦХВЕЯЙНИ ОНЛНЫХ АНКЭМШЛ Б ЮЛАСКЮРНПМШУ СЯКНБХЪУ (НАГНП КХРЕПЮРСПШ).
- пЕГСКЭРЮРШ ЯПЮБМХРЕКЭМНЦН ХГСВЕМХЪ ОЯХУНТХГХНКНЦХВЕЯЙНЦН ЯНЯРНЪМХЪ ЬЙНКЭМХЙНБ Б СЯКНБХЪУ ПЮГКХВМНИ ХМРЕМЯХБМНЯРХ СВЕАМНЦН ОПНЖЕЯЯЮ
- яПЮБМЕМХЕ БНГЛНФМНЯРЕИ ЙНЛОЭЧРЕПМНИ Х ЛЮЦМХРМН-ПЕГНМЮМЯМНИ РНЛНЦПЮТХХ Б НОПЕДЕКЕМХХ УЮПЮЙРЕПХЯРХЙ НОСУНКЕБНЦН СГКЮ Х ЛЕЯРМНИ ПЮЯОПНЯРПЮМ╦ММНЯРХ ПЮЙЮ К╦ЦЙНЦН, НЯКНФМ╦ММНЦН БРНПХВМШЛ БНЯОЮКХРЕКЭМШЛ ОПНЖЕЯЯНЛ
- оСРХ ГЮМНЯЮ ХМТЕЙЖХХ Б БНИЯЙЮ Х ТЮЙРНПШ (СЯКНБХЪ), БКХЪЧЫХЕ МЮ ПЮГБХРХЕ Х ОПНЪБКЕМХЪ ЩОХДЕЛХВЕЯЙНЦН ОПНЖЕЯЯЮ Б ВПЕГБШВЮИМШУ ЯХРСЮЖХЪУ Х Б БНЕММНЕ БПЕЛЪ
- тНПЛХПНБЮМХЕ ЦПСООНБНЦН ОНБЕДЕМХЪ Б НПЦЮМХГЮЖХХ: НЯМНБМШЕ СЯКНБХЪ Х ЯРЮДХХ ЦПСООНБНЦН ОПНЖЕЯЯЮ
- яНЖХЮКЭМН-ЦХЦХЕМХВЕЯЙХЕ СЯКНБХЪ, НОПЕДЕКЪЧЫХЕ НЯНАЕММНЯРХ ДЕЛНЦПЮТХВЕЯЙНИ ЯХРСЮЖХХ Х ОНЙЮГЮРЕКЕИ ГЮАНКЕБЮЕЛНЯРХ ЯЕКЭЯЙНЦН МЮЯЕКЕМХЪ Б ЯНБПЕЛЕММШУ СЯКНБХЪУ
- хГСВЕМХЕ ЩЙНКНЦХВЕЯЙХУ СЯКНБХИ Х ТХГХНКНЦХВЕЯЙХУ ОНЙЮГЮ╜РЕКЕИ ЙКЮПХЕБШУ ЯНЛНБ, ЯНДЕПФЮЫХУЯЪ Б ЮЙБЮПХЮКЭМШУ СЯКНБХЪУ ОПХ ОНДЦНРНБЙЕ Й БНЯОПНХГБНДЯРБС.