3.2. Способы оценки численности носителей и переносчиков возбудителя чумы
Значение численности носителей и переносчиков возбудителя чумы в ее природных очагах трудно переоценить. Сведения о ней используют при оценке эпизоотических, предэпизоотических и любых других ситуаций (Фенюк с соавт., 1963).
Практически всеми исследователями энзоотии чумы признается влияние численности на многие параметры эпизоотических проявлений, включая сами факты их регистрации. Многократно и во многих очагах было установлено предшествование крупным эпизоотиям чумы высокой численности грызунов и блох или, по меньшей мере, повышенных паразитарных контактов на обширных территориях. И наоборот, прекращение эпизоотий или резкое снижение их интенсивности всегда сопровождалось заметным сокращениемi численности животных (Шевченко с соавт., 1976; Денисенко с соавт., 1987; Шевченко, Гражданов, 1989; Улюмджиев с соавт., 1991; Парфёнов, Парфёнова, 2001). Наличие же работ, констатирующих отсутствие зависимости эпизоотической активности от численности носителей и переносчиков, очевидно, следует расценивать как факты упрощенного методологического подхода при решении этого вопроса (Постников, 1983; Руденчик с соавт., 1983; Солдаткин с соавт., 1984; Постников с соавт., 1985; Попов Н.В. с соавт., 1989; 1990; Руденчик, Лубкова, Алексеев, 1989; Руденчик, Солдаткин, Лубкова, 1989; Руденчик, Солдаткин, 1990).Мы не ставили перед собой вопроса — нужно или не нужно определять численность животных, поскольку весь опыт практической работы в очагах чумы и мнение специалистов-чумологов говорят о необходимости этого (Фенюк с соавт., 1963). Проблема заключается в том, как осуществить учет численности быстро, дешево и правильно. Ее решению посвящено большое количество работ, полный обзор которых не входит в нашу задачу, однако, обозначение основной линии в выборе
59 оптимальных методов, которой придерживается автор, привести необходимо.
Для учета численности полуденных и гребенщиковых песчанок был разработан капканно-площадочный метод (Ралль, 1935; 1936 а; Ралль, Демяшев, 1938), применение которого предусматривает вылов грызунов на прямоугольных площадках, закладываемых в типичных местообитаниях. Сначала песчанок учитывали на четвертьгектарных площадках при трех- или двухсуточном их облове. Затем облов был сокращен до одних суток с увеличением размеров площадки до полугектара (Постников, 1955 б). Предлагалось также увеличить площадки до 4-9 га (Солецкий с соавт., 1955) или заменить отлов отравлением зверьков приманкой со стрихнином (Семенов, 1950). В итоге многие годы в противочумной системе применялся односуточный облов одногектарных площадок с предварительной прикопкой нор (Инструкция по учету численности ...,
1978) .
Иной подход к учету численности заключен в отлове грызунов давилками Геро с приманкой, расставленными в линию с определенным интервалом (метод ловушко-линий). Он применяется для учета мелких мышевидных грызунов, хорошо зарекомендовал себя в лесных и кустарниковых биотопах (Кучерук, 1952; 1963) и предлагался также для учета малых песчанок (Бахтигозин, Щепотьев, 1972). Более высокая зависимость результатов учета от активности грызунов, нежели от самой численности (Кучерук с соавт., 1963), ограничивает его применение.
При учете на площадках для обеспечения приемлемой рандомизации проб, необходимо закладывать очень большое количество мест учета и широко рассредоточивать их по территории (Семенов, 1938; Коли, 1979). Однако рандомизация существенно повышается, если учет вести на трансектах большой длины (Лаптев, 1930; 1935; Смирнов, 1976; Коли,
1979) . Маршрутный подход был использован при разработке методов учета больших песчанок, сурков и сусликов (Зверев, Краснов, 1935;
60
Касаткин, 1951; Варшавский, 1952; Лисицын, Миронов, 1953; Бибиков, 1956; 1963; Касаткин с соавт., 1956; Ротшильд, 1957; Касаткин, Стогов, 1963). Учет грызунов-норников, обитающих в открытых ландшафтах природных очагов чумы, облегчается тем, что их жилища (норы, городки, бутаны, курганчики, колонии и т.
д.) хорошо видны на поверхности земли открывающимися на ней норовыми отверстиями. Учет нор видов, ведущих дневной образ жизни, облегчается еще и тем, что их обитаемость визуально подтверждается наличием выходящих из них зверьков (Лаптев, 1930). В большом числе случаев ведут подсчет самих животных.C грызунами, активными ночью, дело обстоит несколько сложнее, хотя определенное мнение об обитаемости нор можно получить по различным следам жизнедеятельности их хозяев, либо вылавливая животных. Маршрутные (линейно-капканные, ленточные и т. д.) методы учета полуденных и гребенщиковых песчанок также стали приходить на смену капканно-площадочному (Бакеев, 1961; Соколов, 1961; Яковлев с соавт., 1964; Павлов, 1972; Ротшильд с соавт., 1975; Бурделов, Тлегенов, Агеев, Шевченко, 1981). Из перечисленных исследователей лишь А.С. Бурделов с соавторами (1981) применили учет двух показателей — количества городков песчанок, встреченных на маршруте, и среднего числа зверьков в одном городке. Положительными моментами в предлагаемых методах служило требование включать различные биотопы в учетную полосу пропорционально их естественному соотношению на той территории, на которую затем будут экстраполироваться данные (Бакеев, 1961), и в качестве объекта визуального учета использовать городок песчанок (Ротшильд с соавт., 1975; Бурделов, Тлегенов, Агеев, Шевченко, 1981).
И при площадочном, и при маршрутном методах учета получают выборку некоторых значений, из которых затем формируют необходимый показатель. Важнейшие свойства всех выборок — их репрезентативность и рандомизированность. Маршрутные методы обеспечивают значительно
61 лучшее качество выборок по этим признакам (Смирнов, 1976) за счет большой длины маршрута. При площадочном методе заложить адекватное их число просто невозможно.
Еще одно свойство любого метода характеризуется уровнем трудозатрат на его осуществление. Например, маршрут с визуальным подсчетом нор может быть пройден одним учетчиком, а в закладке площадок участвуют несколько ловцов.
При учете на площадках результат получают на следующий, а иногда и на третий день. Данные же визуального учета на маршруте обычно готовы сразу после его прохождения.Помимо большой трудоемкости капканно-площадочный метод обладает другими существенными недостатками. Один из них — субъективизм размещения учетных площадок по территории в слабой зависимости от ландшафтной структуры местности. Один из ярких примеров субъективизма — тенденция размещать площадки по выбору (не «случайно») в местах с высокой плотностью нор и зверьков, приводящая к отсутствию площадок с «нулевыми» показателями. При этом всем известно наличие значительных пространств, полностью лишенных нор грызунов. Субъективизм возрастает пропорционально числу площадок, входящих в одну учетную группу. Это же правило (размещение площадок группами) существенно снижает рандомизацию проб в формируемой выборке. Подробную оценку достоверности результатов учета на площадках провел И.П. Бром (1958; 1961; 1963), показавший резкое увеличение ошибки при снижении нормы учета. Приемлемые результаты учета на площадках могут быть получены при равномерном распределении нор по территории (чего в природе не наблюдается), либо при строго равномерном и густом размещении площадок.
Аргументы в пользу маршрутного метода весьма весомы и вполне очевидны. Исходя из этого, был разработан более объективный, логически обоснованный метод оценки численности грызунов — обитателей
62 открытых равнин, названный методом пунктов учета. Вначале он был использован для учета малых песчанок Закавказья (Эйгелис с соавт., 1980; 1982), а затем с необходимыми изменениями — песчанок Волго-Уральских песков (Кузнецов с соавт., 1982; 1983).
Для учета численности полуденных и гребенщиковых песчанок в зоне деятельности Астраханской противочумной станции (ВолгоУральский и Прикаспийский песчаные очаги чумы) этот метод используется уже 25 лет. Результаты учета выражаются привычным показателем численности — усредненной плотностью зверьков, обитающих на 1 гектаре территории очага.
Эта плотность пропорциональна (в пределах статистической ошибки) численности (поголовью) песчанок в зоне экстраполяции. Благодаря большой длине одного маршрута (5 км) и достаточному их количеству, чувствительность и разрешающая способность метода весьма высоки, что особенно четко проявляется в условиях низкой численности песчанок (Кузнецов с соавт., 1982; 1983).При внедрении маршрутного метода для учета песчанок Прикаспийской низменности автором диссертации были уточнены некоторые детали его применения. Объектом учета был избран отдельный (единичный) городок песчанок, размер проекции которого на поверхности земли (при оконтуривании по крайним норовым отверстиям) не превышает 0,01 га (100 м2). В связи с этим учетной единицей на маршруте принят его 10-метровой отрезок при такой же ширине учетного трансекта. Фиксация данных о наличии городков производится на каждом таком отрезке. Иными словами один пятикилометровый маршрут состоит из 500 учетных единиц, — маленьких площадок, составленных в непрерывную ленту. Результаты учета нор на маршруте выражаются долей заселенной территории (в %), вычисляемой по числу 10-метровок, занятых городками, от общего их числа. Эта доля названа «заселенностью территории» и
63 соответствует среднему числу «стандартизованных» городков площадью по 0,01 га на 1 гектаре.
Известно, что в традиционные учетные периоды (апрель и октябрь) в одном городке обитает, как правило, одна взрослая песчанка (Скиртачев, 1985). В идеальном случае число городков должно соответствовать числу песчанок. Однако в реальных условиях бывают периоды, когда число зверьков отличается от числа городков. Например, к апрелю часто происходит существенное снижение численности грызунов, но по внешним признакам городков, которые фактически опустели, этого еще не видно (Постников, 1955 а). Кроме того, возросшая двигательная активность оставшихся зверьков обеспечивает избыток следов на поверхности как жилых, так и нежилых городков. В октябре же наоборот бывает так, что в некоторых городках еще живут с матерью не успевшие расселиться молодые зверьки.
Величина несоответствия количеств песчанок и городков может быть определена при облове городков, который осуществляют на любом удалении от маршрута, но на «подведомственной» ему территории. В каждой местности облавливают не менее 30 городков и при этом определяют среднее число песчанок на один городок стандартной площади (0,01 га), называемое «коэффициентом плотности».За несколько лет ежесезонного облова городков было выяснено, что весной в одном городке в среднем обитает 0,7 песчанки любого вида, а осенью — 1,0. В отдельные нетипичные годы коэффициент плотности мог отличаться от приведенных выше средних значений на 20-30%, однако, мы считаем, что возникающая при этом ошибка в установлении показателя численности не столь высока и ею можно пренебречь. Во всяком случае, выигрыш в производительности труда при учете без облова городков оказывается несоизмеримо выше, чем незначительное снижение точности показателя.
64
Плотность песчанок на 1 га территории определяют перемножением заселенности и сезонного коэффициента плотности. Облов городков в последние годы на Астраханской ПЧС уже не проводится, что чрезвычайно упрощает и ускоряет учетные работы без ощутимого снижения правильности и точности результатов.
Таким образом, в итоге наших исследований методика учета численности полуденных и гребенщиковых песчанок в песках Нижнего Поволжья доведена до наиболее оптимального варианта. Проведение учетных работ описанным способом осуществляется в кратчайшие сроки на всей очаговой территории.
Численности блох стали придавать значение с тех пор, как в природных очагах чумы началось проведение систематического эпизоотологического обследования (Иофф, 1941). Справедливо считалось, что возможность распространения чумы по территории очага напрямую зависит от обилия переносчиков. Разработке методов количественного учета переносчиков в природе придавалось большое значение (Павловский, 1931; Беклемишев, 1931 и другие). Зависимость,, эпидемических проявлений чумы от обилия блох была подтверждена и обоснована И.С.Тинкером (1940), И.Г.Иоффом (1941), В.И.Кузенковым (1941) и другими. Было показано также, что только правильная методика паразитологических сборов обеспечивает получение полноценных материалов для экологических выводов и эпизоотологических прогнозов (Иофф, 1949; Ширанович, 1955).
На первых этапах изучения экологии блох основное внимание обращали на блох гнезда, так как считалось, что именно эта группа эктопаразитов наиболее доступна для получения полных сборов, пригодных для статистической обработки. Однако раскопка нор является очень трудоемким и малодоступным методом для широкого его использования в обыденной практике. Поэтому дальнейшие исследования
65
были направлены на изучение численности блох, обитающих на грызуне. Тем не менее, до определенного времени решающего значения в целях учета численности блох в природе не имел и этот метод (Ширанович, 1955). Как затем было установлено, одним из количественных показателей участия блох в трансмиссивном эпизоотическом процессе является среднее число этих эктопаразитов, питающихся на одном зверьке с бактериемией (Солдаткин, 1963; 1968; Солдаткин, Фенюк, 1968). Предполагалась зависимость этого числа от численности кровососов и от параметров их паразитирования, однако полной ясности в установлении этой зависимости, что наглядно видно по имеющимся публикациям, достигнуто не было. Нам удалось установить эту зависимость в аналитической форме (Кузнецов, 1996; 1997 а), что детально отражено в подразделе 5.1.1. Особое место в установленной зависимости занимает индекс обилия (ИО) блох в шерсти прокормителей, а приемы его правильного определения рассмотрены в данном разделе.
Учет численности блох на хозяине требует соблюдения многих условий для того, чтобы он был достоверен и полноценен. Влияние на величину индекса обилия блох, состояния грызуна в момент заключения его в мешочек, было установлено при изучении эктопаразитов большой песчанки (Федина, Ширанович, 1950). Дальнейший анализ проблемы показал существенную разницу в числе блох на грызунах, добытых живыми или умерщвленными (Ширанович, 1955). Разная скорость ухода эктопаразитов с трупа обусловлена временем суток, видом и размерами зверька, а так же видом блох (Лазарева, Лазарев, 1973; Бурделов с соавт., 1989; Шилов с соавт., 2000). Дополнительным аргументом в пользу более быстрого ухода блох с мелких видов и особей может служить более низкий индекс обилия блох на них (Бурделов с соавт., 1989).
Наиболее ощутимая потеря блох наблюдается у зверьков, активных в ночное время, которые отлавливаются капканами и к моменту съема орудий лова утром оказываются уже окоченевшими, а та или иная часть
66 блох покидает тела своих погибших хозяев. В случае поимки грызуна живым все блохи остаются на нем (по меньшей мере, принято так считать). Поскольку основная масса мелких млекопитающих при эпизоотологическом обследовании (то есть «в рабочем порядке») добывается в умерщвленном состоянии, было предложено разделять индексы обилия блох на «рабочие» и «истинные». Истинные индексы получают по данным очеса зверьков, пойманных живыми (Дойников с соавт., 1955). Указанные авторы выяснили, что истинные индексы в среднем выше рабочих в 2,5 раза на гребенщиковых и в 3,3 раза на полуденных песчанках из Волго-Уральских песков. К сожалению, они не указали размеры выборок, обеспечившие эти результаты.
Позднее аналогичное сравнение индексов было проведено в Прикаспийском песчаном очаге чумы на полуденных песчанках и домовых мышах М.В. Пестовым (1989). Он установил двукратное уменьшение рабочего индекса на песчанках и семикратное — на мышах. Домовые мыши отлавливались только осенью, а полуденные песчанки — весной в относительно небольшом количестве. Близкие результаты были получены при отлове гребенщиковых песчанок в пойме Урала (Агеев, 1974).
Недостаточная репрезентативность данных, полученных нашими предшественниками, не позволяет использовать их в качестве поправок, корректирующих результаты обычных учетов. В СВЯЗИ C этим мы поставили перед собой задачу сравнить ИО блох в шерсти полуденных и гребенщиковых песчанок, пойманных живоловками и капканами, то есть истинные и рабочие индексы. Для этого в 1990 и 1991 гг. на некоторых точках эпизоотологического обследования в Волго-Уральских и Прикаспийских песках добывали грызунов разными орудиями лова. При этом стремились расположить линии тех и других возможно ближе друг к другу, что позволяло считать всех пойманных зверьков обитателями одного поселения с одинаковым исходным числом блох в их шерсти. Мы полагаем, что нам удалось достаточно точно определить степень
67 занижения ИО блох на основных носителях чумы в песчаных очагах Северного и Северо-Западного Прикаспия (Кузнецов, Кабин, Козлова, Синцов, 1996).
При работе на отдельных точках стремились добыть по 50 зверьков одного вида как на линии живоловок, так и давилок. Иногда отлов приходилось вести 2-3 дня для получения выборки необходимой величины. Живых грызунов умерщвляли только после помещения в мешочек и снабжали отдельной этикеткой. Кроме полуденных и гребенщиковых песчанок в достаточном количестве были добыты и домовые мыши, данные о которых также проанализированы.
В таблице 1 приведены данные о добытом материале и результаты сравнения ИО блох отдельно по видам грызунов Волго-Уральских и Прикаспийских песков. В период исследований на песчанках преобладали блохи N. Iaeviceps,на мышах — N. mokrzeckyi.При дроблении материала (вплоть до отдельных точек) отмечены некоторые случайные колебания кратности занижения ИО в шерсти забитых зверьков, однако, не отмечено ни одного случая обратного соотношения, то есть когда на живых зверьках индекс оказывался бы ниже, чем на умерщвленных.
Это весьма красноречивая тенденция, а итоговые цифры (последняя графа таблицы) можно вполне применять как поправки в рабочие индексы, получаемые при эпизоотологическом обследовании очагов чумы. При этом поправочные коэффициенты вполне могут быть округлены до целых значений. Даже в этом случае исправленные индексы обилия будут гораздо правильнее отражать действительность, а возникающие при этом отклонения от истинных значений не будут выходить за рамки допустимой статистической погрешности (Кузнецов, Кабин, Козлова, Синцов, 1996). Так, рабочие индексы обилия блох на полуденных песчанках могут быть увеличены в 4 раза, на гребенщиковых — в 2 и на домовых мышах — в 3 раза.
68
Таблица 1. Соотношение индексов обилия блох на живых и умерщвленных грызунах в 1990-1991 гг.
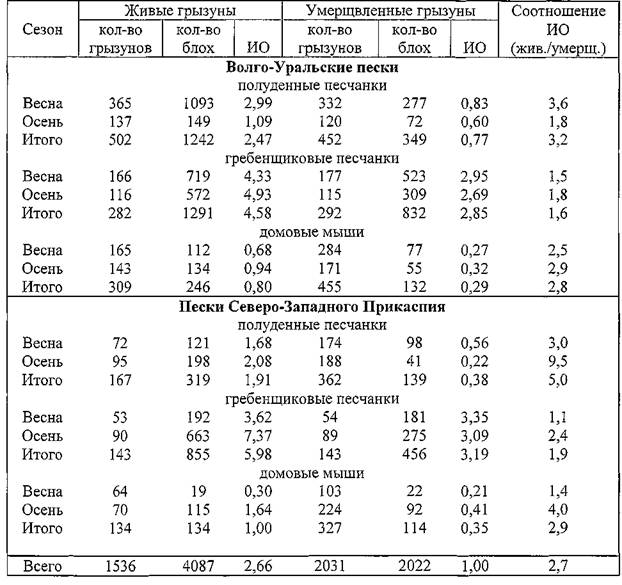
Примечание. ИО — индекс обилия блох в шерсти зверьков.
Знание индекса обилия блох на грызунах необходимо для анализа и оценки условий и возможности развития эпизоотий чумы, в частности — для вычисления коэффициента трансмиссивности. Однако этих данных недостаточно для прогноза дальнейшего состояния популяций переносчиков. Более полная оценка и прогноз возможных изменений эпизоотической активности осуществимы при наличии данных о численности всей популяции блох, точнее, о ее плотности на единицу земной поверхности, то есть «запас» эктопаразитов (Жовтый, 1959).
69
В прошлом «запас» (число блох на 1 га территории или в 1 микробиотопе) определяли путем полной раскопки нор грызунов C выловом всех обитателей и добычей гнезд. Трудоемкость этого мероприятия чрезмерно высока. Отказ от раскопок стал возможен благодаря существованию относительно постоянной приуроченности блох к хозяину, имеющей видовые, популяционные и фенологические особенности. Рядом исследователей была показана возможность применения индекса приуроченности для определения запаса эктопаразитов в микробиотопе (Иофф, 1941; Новокрещенова, 1960; Жовтый, 1960; 1966; Беклемишев, 1961; Старожицкая с соавт., 1978; 1980; Князева с соавт., 1983; Бурделова, Старожицкая, 1986). Количественное выражение приуроченности отражено в индексе, показывающем долю блох, находящихся в некоторый момент времени в шерсти прокормителей, от общего их числа в микробиотопе, поселении или популяции хозяев. При этом особое значение имеет следующая зависимость — число блох, находящихся в шерсти грызунов, относится к их общему числу в микробиотопе (в сумме с находящимися в шерсти) так же, как длительность однократного паразитирования ко всему трофическому циклу. Упоминания о подобной зависимости встречались и раньше (Иофф, 1941; Гаузштейн с соавт., 1965; Солдаткин с соавт., 1967), однако ее математическая интерпретация осуществлена автором диссертации и приведена в подразделе 5.1.1. (Кузнецов, 1996; 1997 а).
Индексы приуроченности блох X. Conformisи N Iaevicepsк полуденным и гребенщиковым песчанкам в песках Прикаспийской низменности были определены путем полной раскопки нор зверьков (Самуров с соавт., 1987; 1990). По данным этих авторов в учетные периоды в шерсти грызунов находится, как правило, не более 10% блох, следовательно, запас блох в микробиотопе достаточно велик. Результаты указанных исследований используются в настоящее время специалистами
70
Астраханской ПЧС при учете численности блох в Волго-Уральском и Прикаспийском песчаных очагах чумы (Эйгелис, Кузнецов, 1989).
Говоря о численности блох нельзя не упомянуть практикующийся способ сбора эктопаразитов из входов нор грызунов. Для этого используются различные приспособления по выемке блох из нор (Ширанович с соавт., 1950; Быков, Белкина, 1964). Однако на основании исследований предыдущих авторов (Колпакова, 1950; Дудникова, 1951; Ворона, 1955; Куницкий, Гаузштейн, 1987) и собственного опыта следует признать, что сборы из входов нор малых песчанок не могут обеспечить массового паразитологического материала для лабораторного исследования. Определение индексов обилия мигрирующих блох для целей учета численности не требуется, да и активность их нападений на человека в песчаночьих очагах достаточно низка (Кузнецов с соавт., 1997). Единственная необходимость осмотра входов нор заключена в индикационном значении усиленной миграции блох к устьям по причине гибели прокормителей. Такая миграция, например, может служить наводящим признаком при поиске мест протекания эпизоотий чумы.
Данные по специальному очесу живых зверьков показали, что истинные индексы обилия блох на полуденных и гребенщиковых песчанках в обоих очагах ни разу не были меньше 1.0. При изучении разноса блох на стационарных участках наблюдений (СУН) в ВолгоУральских песках средние ИО также значительно превышали 1.0. При этом колебания числа счесываемых блох с одного и того же зверька в разные дни были весьма значительными — от 0 до 26. Результаты очесов живых песчанок, регулярно отлавливаемых на СУН, могут служить источником дополнительной информации о численности блох (Агеев, 1974). В нашей работе, осуществляя многократные очесы зверьков, можно было рано или поздно ожидать снижения индекса блох в шерсти меченых песчанок за счет изъятия большей части эктопаразитов из микробиотопов, составляющих данное поселение зверьков. Однако этого обычно не
71
происходило, за исключением весны 1994 г., когда было отмечено резкое снижение численности блох N. Iaeviceps.Во второй половине мая произошло естественное и закономерное отмирание имаго этих блох, совпавшее с нашими работами. Динамика индексов обилия (ИО) блох на некоторых меченых грызунах приведена в таблице 2, из которой отмеченное выше отмирание можно видеть по очесам гребенщиковых песчанок №№ 44, 45 и 54. В другие периоды никаких закономерных изменений ИО нами отмечено не было. Регистрировались только хаотические и порой резкие его колебания. Данные таблицы 2 служат наглядной иллюстрацией этому. Для имитации «изъятия» блох в те сезоны, когда их метили и выпускали, все повторно обнаруживаемые меченые эктопаразиты не учитывались при вычислении индекса. Данные о «постоянстве» индексов обилия блох в шерсти и о его высоком среднем уровне свидетельствуют о значительном запасе эктопаразитов в поселениях песчанок. Эти данные, наряду с результатами изучения форезии блох, говорят о более существенной «насыщенности» природного очага чумы переносчиками, чем это можно видеть по результатам обычного обследования. Кроме того, постоянные переносы большого количества блох из норы в нору лишают определенности понятие «микропопуляция» (Беклемишев, 1959) и наполняют более глубоким смыслом понятие «популяция» блох. Очевидно, пространственные параметры популяций эктопаразитов тождественны подобным параметрам популяций их хозяев.
Таким образом, мы установили, что данные об индексах обилия блох, получаемые при плановом эпизоотологическом обследовании, существенно занижены и должны быть увеличены в 2-4 раза. Для углубленного научного и эпизоотологического анализа, включающего вычисление коэффициента трансмиссивности чумы, необходимо в результаты обследования регулярно вносить поправку.
Таблица 2. Динамика индекса обилия блох в шерсти живых песчанок при их многократных очесах в 1990-1994 гг. на стационарных участках наблюдений в Волго-Уральских песках

Примечание. В скобках указано число повторно обнаруженных меченых блох, которые не учитывались при вычислении индекса обилия.
73
3.3.
Еще по теме 3.2. Способы оценки численности носителей и переносчиков возбудителя чумы:
- Структура поселений и численность носителей и переносчиков возбудителя чумы на плато Тайнгуен
- Оценка передвижений и контактов носителей и переносчиков возбудителя чумы.
- Оценка трансмиссивного пути передачи возбудителя чумы и структура коэффициента трансмиссивности
- Блохи как переносчики чумы.
- Оценка активности природных очагов чумы — основа их пространственной дифференциации.
- Оценка биоценотической и пространственной структуры очагов чумы
- 2.5 Методология оценки чувствительности возбудителей онихомикоза к антимикотическим препаратам
- 1.2. Способы оценки двигательных нарушений.
- 4.1. Способы оценки косметического результата лечения.
- Графические способы оценки КМП
- Способы оценки эффективности работы профконсультантов.
- 21. Способы оценки условий и приёмов выдвижения лидеров группой.
- Мембранные переносчики.
- Составление обзора и прогноза численности эктопаразитов
- Вероятность наступления бактериемии у носителей