Перемещения полуденных и гребенщиковых песчанок в Волго-Уральских песках.
После того, как М.М. Тихомировой (1936) в 1926-1933 гг. была установлена ведущая роль полуденной песчанки в поддержании энзоотии чумы в Волго-Уральских песках, началось целенаправленное изучение всех сторон жизнедеятельности этих и других пустынных грызунов (Ралль, 1938; 1939 а; 1939 б; 1941).
Среди различных вопросов изучались миграции и прочие формы передвижений песчанок методом кольцевания (Ралль, 1936 а; 1936 б; Фенюк, Демяшев, 1936 а; 1936 б), уже успешно применявшимся для мечения сусликов (Зверев, 1927; Калабухов, 1933; Калабухов, Раевский, 1935). Работа Б.К.Фенюка и М.П.Демяшева (1936 б) по мечению полуденных и гребенщиковых песчанок, проведенная в 1933— 1935 гг., является одним из примеров классического подхода к решению проблемы. Авторы установили, что дальние массовые миграции не характерны для этих грызунов. Вместе со значительной привязанностью к своим местообитаниям они обладают весьма высокой подвижностью, что подтверждается и другими исследователями (Ралль, 1931; 1938; 1941). По данным Б.К.Фенюка и М.П.Демяшева (1936 б) дальность перемещений большинства зверьков ограничивалась сотней метров, а предельное зафиксированное расстояние, на которое переселилась одна полуденная песчанка, составило 1 км. Позднее другими исследователями были установлены отдельные факты наибольших перемещений полуденных и гребенщиковых песчанок за тот или иной срок. Обычно предельные расстояния миграций этих грызунов ограничивались 400-800 м, порой достигая 1.5 км, а в некоторых случаях превышали 3 км (Бахтигозин с соавт., 1961; Тропин, 1965; Медзыховский, 1971 б; Медзыховский, Маштаков, 1974). Лишь в одной работе были зафиксированы весьма небольшие перемещения полуденных песчанок (Ефимов с соавт., 1989), что, возможно, было связано с несовершенством использованной методики.' 245
Все эти расстояния были измерены авторами при отлове меченых зверьков по прошествии нескольких дней и даже месяцев после мечения.
В одном случае было отмечено перемещение гребенщиковой песчанки на 571 м за одну ночь (Медзыховский, 1971 б). Отмечено несколько случаев быстрого (через небольшое число дней) переселения на 2-3 км, происходившего обычно ранней весной (Медзыховский, Маштаков, 1974). Некоторое представление о перемещениях песчанок Волго-Уральских песков можно получить также по результатам разноса меченых блох из работы Н.А.Мокриевича с соавт. (1969). Авторы всех упоминавшихся работ отмечали сочетание у малых песчанок высокой повседневной подвижности с общей оседлостью. Некоторые зверьки все же имели склонность к постоянным миграциям, хотя, как нам кажется, это явление осталось слабо изученным.Полноценная характеристика подвижности носителей может быть получена при оценке реальных ежесуточных перемещений с возможно более частой регистрацией зверьков. Попытки измерить средние расстояния, которые преодолевают грызуны за сутки, предпринимались и раньше. Так, Б.К. Фенюк и М.П. Демяшев (1936 б) определяли «среднесуточную скорость передвижения» делением дистанции, разделяющей пункты последовательных поимок зверька, на число дней между этими поимками. Прием расчета, применявшийся в те годы, приводил к неадекватности отдельных результатов, составлявших порой лишь доли метра. Такие результаты получались, когда между поимками зверька проходил длительный срок, а расстояние между пунктами находок оказывалось таким же обычным, как при вылове зверька через небольшое число дней или даже меньшим.
Более реальное значение среднего расстояния, на которое перемещается зверек за сутки, приведено в статье Г.А. Медзыховского (1971 б) — весной оно было равно 63,1 м, а осенью — 19,0 м. К сожалению, автор не привел способа вычисления и объема данных,
246 использованных для этого. Мы в своей работе применили собственную методику количественных оценок (Кузнецов, Матросов, 2004), о чем будет сказано при обсуждении результатов.
Перемещаясь, грызуны помимо функций собственного размножения и расселения обеспечивают осуществление этих же функций своим эктопаразитам.
Обмен блохами возможен без непосредственной встречи донора и реципиента — достаточно посещения одной и той же норы в разные моменты времени, а сам факт смены хозяина означает реализацию паразитарного контакта между ними. Хотя было показано, что переход блох с одного зверька на другого может происходить и при их непосредственном контакте (Краснов, Князева, 1983), роль его незначительна. Поскольку опосредованные паразитарные контакты происходят в норах, то и расстояния перемещений зверьков следует учитывать в случаях фактического достижения ими реальных нор, а не абстрактных точек пространства. Это положение определило в нашей работе расстановку орудий лова только возле нор на участке наблюдений.Продолжительность транспортировки блох хозяином сопоставима с длительностью их непрерывного паразитирования и обычно не превышает одних суток (Кузнецов, 1997 а). В связи с этим для оценки дистанций между пунктами возможных паразитарных контактов особую важность приобретает определение средних расстояний перемещений зверьков в течение суток. Тем не менее, и суточные, и более длительные перемещения обеспечивают осуществление социальных контактов, необходимых для поддержания внутрипопуляционного гомеостаза.
На начальном этапе исследований в процессе поиска и отработки необходимых методик планировалось использовать в качестве основного метода визуальные наблюдения за маркированными зверьками со специально построенной для этого вышки, так же, как наблюдали, например, за большой песчанкой (Панченко с соавт., 1971). В ночное время такие наблюдения вели при искусственном освещении площадки
IAI (Веревкин, Миронов, 1983). Мы полагали, что с помощью этой методики могла быть получена информация не только о дальности ежесуточных перемещений полуденных и гребенщиковых песчанок, но и о том, какие норы посещают зверьки, сколько времени проводят в них, а также прочие детали их суточной активности. Эти данные предполагалось использовать для вычисления вероятности смены прокормителя блохой по алгоритму, описанному в подразделе 5.1.3.
Для реализации этого плана нами были выбраны стационарные участки наблюдений (СУН) в центральной части Волго-Уральских песков. Такие места были выбраны как удовлетворяющие ряду условий. Во- первых, площадка должна была находиться в непосредственной близости от помещения, где жили бы экспериментаторы и могла быть организована лаборатория по ежедневному очесу грызунов, определению и мечению эктопаразитов. Во-вторых, необходима была уверенность в том, что в каждый следующий сезон на площадке будут обитать в достаточном количестве полуденные и гребенщиковые песчанки. При этом исключались как зависимость от автотранспорта, так и перспектива вынужденного переноса площадки, в результате которого могла быть нарушена преемственность информации от сезона к сезону. Таким условиям отвечали два места — урочища «Зулхаш» и «Новая Школа» на территории Таутюбинского эпид отряд а, выставляемого Досангским отделением Астраханской противочумной станции. В этих урочищах имелись стойкие поселения обоих видов малых песчанок. Устойчивость конкретно этих поселений была установлена в процессе многократного их картографирования в предшествующие 5 лет (Кузнецов с соавт., 1989; 1991).
C 1989 по 1991 г. (шесть сезонов) работы проводились в урочище «Зулхаш», расположенном в Гурьевской (Атырауской) области Казахстана, а с весны 1993 г. (после распада СССР) по весну 1994 г. (три сезона) — в
25 км западнее предыдущего участка в урочище «Новая Школа» (Астраханская обл.).
В урочище «Зулхаш» площадь СУН составляла в первые сезоны 3-4, затем — 8 га. Лишь весной 1989 г. пробный отлов песчанок был осуществлен на площади 1 га. Участок был расположен среди бугристых песков, заросших песчаной полынью, кустами джузгуна, а в промежутках между ними — эфемеровым разнотравьем. Рельеф участка достаточно сложный с перепадами высот более 4,5 м. Крутизна склонов в отдельных местах достигала 25 угловых градусов. Эта информация получена при помощи теодолитной съемки рельефа, нор и т. д.
Для съемки на местности готовили долговременные точки съемочного обоснования в виде врытых в землю низких и высоких столбиков (Господинов, Сорокин, 1974; Руководство по топографической съемке ..., 1977).
Два низких столбика в центре участка, между которыми было точно измерено мерной лентой базисное расстояние, служили для установки над ними теодолита. Местоположение нор и живоловок определялось угловыми засечками при помощи теодолита и мерной рейки с привязкой к точкам съемочного обоснования (Гараевская, Малюсова, 1976; Ганьшин с соавт., 1977). В результате этой работы появлялась возможность точного и быстрого измерения расстояний между любыми объектами, имеющимися на участке и нанесенными на план. На первом участке применили сгущение сети съемочного обоснования для съемки рельефа местности.В урочище «Новая Школа» площадь СУН составляла около 9 га. Рельеф и растительность на нем имели такой же характер, как и в урочище «Зулхаш». Однако специальную съемку рельефа здесь не проводили, так как не планировали визуальных наблюдений с вышки.
Весной 1989 г. мы предприняли попытку провести визуальные наблюдения за перемещениями песчанок в ночное время. Для этого в центре участка на высоком бугре построили наблюдательную вышку
249 высотой около 4 м. На некотором расстоянии от нее (10-20 м) расставили 5 осветительных установок с лампами, мощностью по 200 Вт. Электропитание ламп осуществлялось при помощи бензоэлектроагрегата, мощностью 1 кВт, расположенного в 200 м от СУН.
Наблюдения с вышки вели с помощью 12-кратного бинокля. При этом сразу же обнаружилось, что характер рельефа и произрастающей растительности на участке чрезвычайно затрудняют прослеживание путей передвижения зверьков по поверхности земли даже днем. По этой причине нам ни разу не удалось сколько-нибудь долго наблюдать конкретных особей, хотя отдельные фрагменты их перебежек были замечены. При наличии всего лишь одного наблюдателя моменты выхода животного из норы или ухода в нее не были зафиксированы ни разу, несмотря на довольно большое число нор на участке. Тем не менее, утром при спуске с вышки обнаруживалась значительная заслеженность незадернованных участков песчанками.
Попытки визуальных наблюдений были продолжены осенью 1989 г.
и весной 1990 г. с аналогичным результатом, после чего от этого метода пришлось отказаться. Имеющийся у нас опыт изучения ландшафтных особенностей Волго-Уральских песков и дислокации поселений малых песчанок в них (Матросов с соавт., 1987; Кузнецов с соавт., 1989; 1990; 1991; Матросов, Кузнецов, Ерофеев, 1996; Кузнецов, Матросов, 2004), убеждает в том, что в этом регионе практически отсутствует возможность подобных визуальных наблюдений за этими грызунами. Можно было бы найти участки голых барханов со сглаженным рельефом, где встречаются поселения полуденных песчанок (но без гребенщиковых), и где, вероятно, удалось бы пронаблюдать за их перемещениями от норы к норе, но на таких участках поселения не сохраняются долго на одном и том же месте и, порой, отсутствуют по несколько сезонов подряд. В данном случае не соблюдаются главные условия, предъявляемые к СУН и изложенные в начале раздела.Таким образом, главным методом изучения подвижности малых песчанок был избран метод многократных отловов зверьков живоловками, который мы начали применять с первых дней работы, сначала как дополнительный, затем как основной. Этот метод давно используется для изучения экологии грызунов (Кучерук, 1963; Никитина, 1964; 1965; 1972; Тропин, 1965; Литвин с соавт., 1973; Попов С.В. с соавт., 1989; Соколов с соавт., 1989), а нами были отработаны лишь детали его применения, необходимые для достижения намеченной цели. В начале каждого сезона по результатам теодолитной съемки вычерчивали план дислокации на участке нор, живоловок и т. д., что давало возможность точного измерения расстояний между любыми объектами, нанесенными на план. Для регистрации перемещений зверьков использовали их индивидуальное мечение ампутацией пальцев (Калабухов, 1951; Кучерук, 1952; Никитина, 1980; Карасева, Телицина, 1996) с последующими многократными отловами ловушками с приманкой. Использовали деревянную живоловку Зайцева (Бурделов, 1951), хорошо зарекомендовавшую себя при работе в различных температурных режимах. Орудия лова расставляли возле нор в 10-20 см от наиболее используемых песчанками входов.
Принимая во внимание объективно ограниченные возможности метода многократных отловов, мы могли изучить с его помощью важный для нашего исследования вопрос — норы каких соседей способна посетить каждая песчанка при ежесуточных передвижениях по своему участку обитания и как далеко расположены эти норы от собственного жилища конкретного зверька. Для этого требовалось расставлять живоловки только у нор грызунов. При этом предполагалось, что если зверек был пойман у какой-либо норы, то он мог либо заходить в эту нору до поимки в качестве посетителя, либо выйти из нее после дневки в качестве хозяина. Учитывая возможность поимки около городка не только его хозяина, но и гостя, стремились выставлять по 2 ловушки на каждый городок в непосредственной близости от наиболее используемых зверьками норовых
251 отверстий (при достаточном количестве орудий лова). Данные о вылове зверьков вдали от нор (если бы там стояли живоловки) были бы не информативны по признаку осуществления норовых контактов, а сведения об абстрактной величине индивидуальных участков нам не требовались.
В первые годы работы на участке «Зулхаш» живоловки, стоящие у нор постоянно, заряжались ежедневно, а в отдельные сезоны — дважды в день, утром и вечером. Это делалось для уточнения периодов активности зверьков. Кроме того, проводили ежедневный очес максимального количества зверьков с целью изучения динамики индекса обилия блох в их шерсти.
Новой деталью методики, которую применили в 1993-1994 гг., явилась оригинальная очередность перезарядки ловушек, выставлявшихся возле нор, как и раньше стационарно. Если живоловка заряжалась каждую ночь, в нее зачастую также каждую ночь попадалась песчанка, обитающая в данной норе. При этом исключалась не только возможность ее перемещений по участку, но и возможность поимки в эту ловушку прибегающих со стороны зверьков. Эту проблему решили следующим образом. Разбили (условно) участок на 4 части с приблизительно равным количеством живоловок в каждой. В первый день работы заряжали все живоловки. При этом удавалось поймать и пометить значительную часть обитавших на участке зверьков. В последующие три дня ни одну из сработавших ловушек не перезаряжали, а грызунов продолжали ловить в остающиеся настороженными орудия лова до завершения четырехдневного цикла. После этого вновь заряжали все живоловки, но только на одной части СУН (первой), на следующий день - на другой части и так далее, по кругу. При таком способе «высокоуловистые» орудия лова автоматически оказывались заряженными лишь одну ночь из четырех, составляющих цикл, менее уловистые — 2-3 ночи, а самые «низкоуловистые» стояли заряженными полный цикл и более. Приманка в заряженных ловушках ежедневно заменялась на свежую. К концу работы
252 практически ни одна живоловка не оставалась не сработавшей хотя бы раз. Иногда, при быстром «расходовании» заряженных ловушек, цикл сокращали до 3 или 2 дней. В случае слишком долгого бездействия ловушку переставляли на новое место. Таким образом, предложенный нами прием работы исключал прикормку зверьков к определенным живоловкам и достаточно быстро выявлял их потенциал подвижности.
Пойманных рано утром грызунов метили, если проводились паразитологические исследования, очесывали, а затем отпускали на месте поимки. Записывали в журнал всю информацию о фактах поимок зверьков — дату и номер живоловки, вид, пол и номер грызуна, количество, виды и номера блох на нем и т. д. Эту работу вели ежедневно на протяжении всего отпущенного на нее срока (25-50 дней в сезон). При обработке полученной информации ежесезонно вычерчивали по данным теодолитной съемки план дислокации живоловок на участке. Используя данные многократных отловов грызунов, также каждый сезон составляли таблицу с датами и расстояниями между местами поимок. Информация, полученная таким способом, позволила достаточно подробно охарактеризовать подвижность песчанок в повседневном плане и ее роль в разносе эктопаразитов по территории, занятой их поселениями.
Общее число городков песчанок обоих видов на каждом из участков не превышало 50. В различные периоды выставляли от 45 до 70 живоловок в соответствии с расположением и числом имеющихся на участке нор. Расстояния между ловушками в пределах одного городка составляли 3-6, разных — 8 и более метров. Дистанция между орудиями лова, стоящими на противоположных краях площадки, достигала 250-300 м.
Особенностью примененного метода является то, что отлов грызуна искусственно ограничивает свободу его дальнейших перемещений вплоть до момента выпуска на волю. Поэтому расстояния между пунктами очередных поимок обычно занижены. Кроме того, при последовательных обнаружениях зверька в одной и той же ловушке возникают «нулевые»
253 результаты, которые мы не учитывали, поскольку факты «неперемещения» на самом деле таковыми не являются.
За период работы на первом участке («Зулхаш») было помечено 137, на втором («Новая Школа») — 11 полуденных песчанок, большинство из которых (86 и 8 соответственно) попадались от 2 до 24 раз, остальные (51 и 3) — только 1 раз при мечении. Гребенщиковых песчанок помечено на первом участке 75, на втором — 66, из них соответственно 57 и 32 зверька попадались от 2 до 21 раза, остальные (21 и 14) — однократно. Среди меченых зверьков большую часть составляли самцы — 62.2% у полуденных и 66.9% у гребенщиковых песчанок.
За весь период работы (без учета «перемещений» от сезона к сезону) измерено 252 перемещения полуденных песчанок, совершенных 95 зверьками. Отдельные перемещения колебались от нескольких метров (в пределах одного или двух соседних городков) до 263, среднее расстояние составило 53.2 м. За сутки по данным 77 измерений песчанки удалялись от места предыдущей поимки в среднем на 41.6 м (максимум на 222), а при более длительном интервале (175 измерений) - на 58.2 м.
При измерении 412 перемещений (также без учета «межсезонных»), совершенных 109 гребенщиковыми песчанками, их расстояния составили от нескольких до 250 м, среднее - 56.8 м. За сутки (153 измерения) зверьки удалялись в среднем на 46.8 м (максимум на 237), за большой срок (259 измерений) - на 62.7 м. Похожие показатели подвижности этих видов песчанок известны из других частей их ареалов (Крючков с соавт., 1957; Агеев, Трофимов, 1974; Тлегенов с соавт., 1974; Карулин с соавт., 1979; Попов, 1981; Веревкин, Миронов, 1983; 1989).
Интервалы времени между двумя смежными поимками песчанок колебались от 1 суток до нескольких недель и месяцев. Продолжительность регистрации зверька на участке (между первой и последней поимками) в пределах одного сезона часто достигала 40-50 дней, а некоторые зверьки (8 полуденных и 12 гребенщиковых песчанок) обитали на участке по 2 сезона
в течение 140-225 дней. При этом лето пережили 1 полуденная и 3 гребенщиковые песчанки, зиму — 7 и 10 зверьков соответственно. Любопытно, что, пережив лето 1993 г. самец гребенщиковой песчанки № 7 в период осенних работ не был пойман ни разу, а следующей весной (пережив еще и зиму) вновь отлавливался на участке. В общей сложности этот грызун был пойман 9 раз в течение 328 дней.
Изменение мест поимок зверьков, доживших до следующего сезона, было соизмеримо с обычными ежесуточными перемещениями. Полуденные песчанки «передвинулись» в среднем на 76.2 м (от 9 до 197), гребенщиковые — на 62.8 м (от 0 до 220), а затем продолжали встречаться практически на тех же или примыкающих к ним участках, что и в прошлый период. Ни один из этих случаев считать настоящим переселением нет оснований, а если таковое происходило, то за пределы участка наблюдений и не могло быть зарегистрировано нами. Возможно отсутствие самца № 7 осенью 1993 г. означало его переселение (выселение), а весной 1994 г. он вернулся на участок. В любом случае «межсезонные» передвижки для расчета подвижности не использовались.
В мае 1989 г. в центральной части СУН «Зулхаш» на площади около
1 га пробный отлов песчанок вели в течение 3-х ночей для определения наличия и численности обитавших на нем грызунов. Было поймано только 3 гребенщиковые песчанки, активность которых была явно снижена. Дело в том, что за зиму 1988-1989 гг. произошло чрезвычайно резкое снижение поголовья песчанок после аномально высокой их численности осенью 1988 г. (в среднем — 60 особей на 1 га по всему юго-западу Волго-Уральских песков). Это обстоятельство совместно с ранним наступлением летней жары обусловили нетипичную («выжидательную») пассивность пустынных грызунов, причем отрицательное влияние «кризисной» зимы сказалось прежде всего на полуденных песчанках, что отмечалось и ранее (Тропин, 1963). Кроме того, наши основные усилия в этот сезон были направлены на попытки ночных визуальных наблюдений, не давших, к
255 сожалению, положительных результатов, поэтому данные о повторных отловах за этот сезон отсутствуют.
В октябре 1989 г. было уделено большее внимание работе по мечению и многократному отлову песчанок живоловками. В это время было поймано и помечено 16 полуденных и 15 гребенщиковых песчанок на площади 3 га. Некоторые зверьки встречались лишь однажды (при мечении), что позволяет считать их «мигрантами», заходившими на площадку со стороны. Для расчета численности постоянных обитателей площадки, мигрантов не учитывали, следовательно, 7 из 16 полуденных песчанок были мигрантами, а 9 — постоянно обитали на площадке (3 особи на 1 га). Из 15 гребенщиковых песчанок постоянно обитали 11 зверьков (3,7 особей на 1 га).
В дальнейшем был увеличен акцент на изучение подвижности методом многократных отловов, а с октября 1990 г. полностью перешли на этот метод. Площадь СУН также увеличивалась — до 4 га в 1990 и 8 га в 1991 г. Позднее (в 1993 г.) в связи с распадом СССР, пришлось изменить дислокацию участка наблюдений, переместив его из Казахстана в Астраханскую область на 25 км западнее. Здесь также имелось стационарное помещение, пригодное для организации лаборатории, и отмечались достаточно устойчивые поселения песчанок. На этом участке («Новая Школа») наблюдения за перемещениями зверьков вели на площади 9 га в 1993—1994 гг.
Численность песчанок непосредственно на площадках мечения во все сезоны держалась несколько ниже обычного среднего уровня, который по сумме видов держится в пределах от 5 до 15 особей на 1 га (Ралль, 1935; 1938; Ралль, Демяшев, 1938; Семенов с соавт., 1968; Павлов с соавт., 1977; Тлегенов с соавт., 1981; Кузнецов с соавт., 1982; Булычев с соавт., 2001). Данные об изменениях численности (плотности) грызунов на стационарных участках наблюдений представлены на рисунке 22.
256
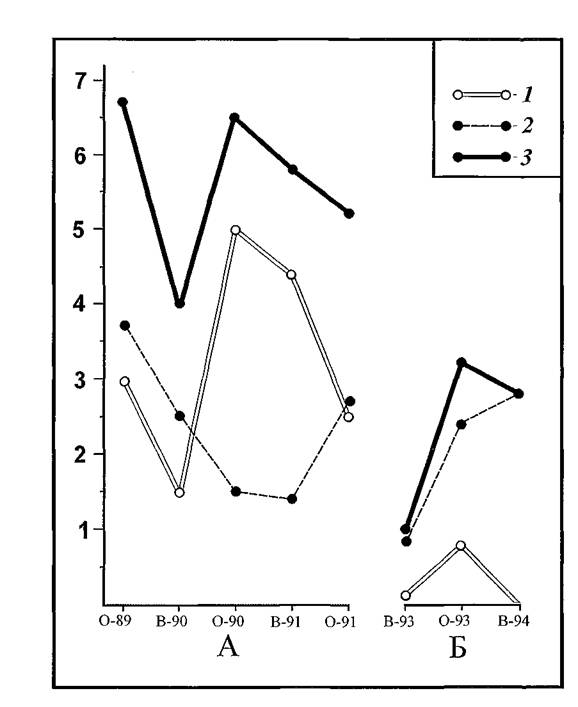
Рис.22. Динамика численности полуденных (1) и гребенщиковых (2) песчанок на стационарных участках наблюдений «Зулхаш» (А) и «Новая Школа» (Б) в 1989-1994 гг. в Волго-Уральских песках. На оси абсцисс — периоды исследований, на оси ординат — число особей на 1 га; 3 - сумма видов.
Следует заметить, что полноценная и правильная оценка численности грызунов должна содержать в себе данные о количестве мигрантов (Щипанов, 1990), однако, при этом трудно обойтись без некоторых допущений. Например, известно, что каждая конкретная особь не является изначально и постоянно либо кочующим, либо оседлым животным, в связи, с чем большинство зверьков могут не раз в течение жизни менять статус оседлости. В нашей работе мы условно выделили группу мигрантов, попавшихся в ловушку только один раз. Условность заключается в том, что некоторые оседлые зверьки крайне осторожны (особенно беременные самки) и редко заходят в живоловки. Среди однократно встреченных грызунов могли быть и такие самки. C другой стороны, некоторые особо подвижные «бродяги» в силу повышенной активности могли неоднократно пересекать площадку мечения и быть зачисленными в группу оседлых. Однако мы надеемся, что такие варианты были редки и не могли серьезно повлиять на общую оценку численности.
Данные о перемещениях формировались в тех случаях, когда меченые песчанки обнаруживались в других ловушках относительно предыдущих поимок и когда могли быть измерены расстояния перемещений (разумеется, условные — «по прямой» между местами отлова). Фактически грызун мог двигаться по совершенно иному маршруту, однако мы не имели возможности измерять ничего другого, кроме расстояния между живоловками. Если зверек в очередной раз попадался в ту же ловушку, он лишался возможности двигаться и такие факты «неперемещения» мы просто не учитывали. Анализу подвергались данные не о количестве поимок как таковых, а измеренные расстояния между местами очередных поимок.
В соответствии с приведенными выше уточнениями в таблице 18 приведены итоговые данные и параметры перемещений самок и самцов полуденных и гребенщиковых песчанок за все годы и сезоны исследований. Анализ данных по отдельным группам показывает
Таблица 18. Сводные данные о подвижности самок и самцов полуденных и гребенщиковых песчанок в Волго-Уральских песках в весенние и осенние сезоны 1989-1994 гг.
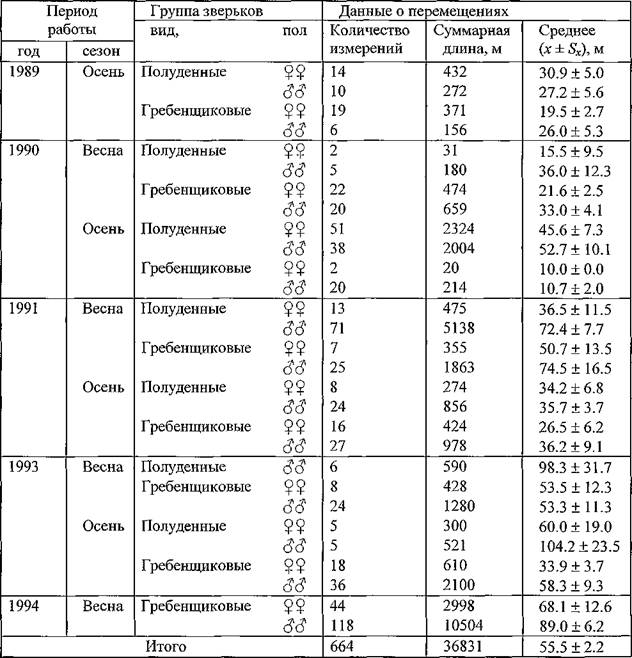
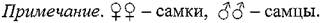
существенную вариабельность средних значений. Достаточно устойчиво показатели подвижности самцов обоих видов песчанок превышали показатели самок. Для полноценного сравнения и выявления различий между полами были пригодны вариационные ряды перемещений гребенщиковых песчанок за весну 1990 и 1994 гг., весну и осень 1991 и
1993 гг. (6 сезонов), полуденных песчанок — за осень 1990 г., весну и осень 1991 г. (3 сезона).
При сравнении средних показателей подвижности самок и самцов гребенщиковых песчанок отмечено наложение диапазонов ошибок репрезентативности в оба сезона 1991 г. и весной 1993 г. (таблица 19). Диапазоны не достигали друг друга весной 1990 и 1994 гг., а также осенью 1993 г. Имело смысл проверить достоверность лишь этих, заметных на глаз различий. Проверка значимости различий по Х-критерию Ван-дер- Вардена (Лакин, 1990) показала сомнительный результат (0.01
Еще по теме Перемещения полуденных и гребенщиковых песчанок в Волго-Уральских песках.:
- Форезия блох в Волго-Уральских песках
- Ландшафтная приуроченность элементов пространственной структуры популяций песчанок в ВолгоУральском песчаном очаге чумы.
- Волго-Уральский песчаный очаг(16)
- Структура Волго-Уральского песчаного очага чумы.
- Ротационные перемещения.
- Поперечные перемещения отломков
- Параграф десятый. Признаки кризиса путем перемещения
- Исследование стабилометрических характеристик произвольных целенаправленных перемещений центра давления
- Угловые перемещения,
- Параграф двенадцатый. Признаки перемещения в верхние, части тела
- Патологические процессы, вызываемые перемещением по геном; человека LI*
- Патологические процессы, вызываемые перемещением по геному человека Alu-элементов*
- Перемещения малых крыс в сельских населенных пунктах на плато Тайнгуен
- Параграф одиннадцатый. Признак того, что перемещение направлено в нижние части тела
- ВЫВОДЫ