Форезия блох в Волго-Уральских песках
Блохи — паразитические насекомые, имаго которых являются облигатными гематофагами. Значительная часть видов блох обнаруживает смешение признаков гнездово-норовых и временных эктопаразитов с кратковременным питанием (Беклемишев, 1945; 1954; 1972; Балашов, 1982).
Виды, являющиеся переносчиками возбудителя чумы, в течение жизни регулярно покидают тело хозяина во время перерывов между периодами кровососаний, затем нападают на него вновь. Находясь в шерсти грызуна-прокормителя, блоха перемещается вместе с ним по территории и имеет возможность покинуть его не в той норе, где было совершено нападение. Покидая покровы тела одного хозяина, блоха получает возможность в следующий раз напасть на другого. Транспортировка блохи грызуном, сопутствующая периодам паразитирования, носит название форезии, а подобный вид биоценологической связи назван прямой подчиненной форической связью (Беклемишев, 1951; 1972).Изучение форезии, или разноса блох грызунами необходимо для количественной оценки чумного эпизоотического процесса. Одним из условий, определяющих возможность передачи возбудителя чумы от грызуна грызуну, является многократное чередование периодов паразитирования и свободного существования блох-переносчиков в норах. Временное пребывание зараженного эктопаразита в шерстном покрове перемещающегося по поверхности земли прокормителя, уход кровососа в одной из посещенных зверьком нор, а затем очередное нападение на другого животного, оказавшегося в норе, — все это обеспечивает возможность развития и территориального распространения эпизоотии чумы. Важными характеристиками процесса форезии блох, подлежащими изучению, служат расстояния переноса эктопаразитов из норы в нору и вероятность смены ими прокормителей.
Масштабы форезии блох, на наш взгляд, не были изучены с достаточной подробностью, несмотря на регулярно предпринимавшиеся для этого усилия.
Объективное изучение разноса блох возможно при помощи их мечения, в отличие от анализа данных о паразитировании неспецифических видов (Миронов с соавт., 1949; Ширанович, Миронов, 1956; Шевченко с соавт., 1965). Предыдущими исследователями использовалось лишь массовое мечение насекомых либо красителями (Мошковский, Носина, 1933; Косминский, Соловьева, 1959; Ширанович с соавт., 1959; Деревянченко с соавт., 1960; Никитина, Николаева, 1980), либо радиоактивными изотопами (Новокрещенова с соавт., 1961; Солдаткин с соавт, 1961; 1962; Руденчик, 1963; Морозова с соавт., 1969; Солдаткин, Руденчик, Корнеев, Северова, 1969; Солдаткин, Руденчик, Северова, 1969 и многие другие). Параллельно этим методам применяли ампутацию одной из лапок блохи (Косминский, 1960; Дудникова с соавт., 1964; Лукьянова, Лапина, 1965; Лапина с соавт., 1968), однако, и в этом случае мечение не было индивидуальным. Кроме того, все перечисленные способы требовали безвозвратного изъятия насекомых из природы для однократной регистрации их перемещений за время, прошедшее после мечения.Метод ампутации члеников лапок не получил широкого распространения, однако, привлек наше внимание как перспективный для усовершенствования. Разработанный на его основе прием обеспечивает присвоение каждой блохе индивидуального номера, распознавание которого не связано с умерщвлением и изъятием насекомого из природы (Кузнецов с соавт., 1993; Кузнецов, Матросов, Козлова с соавт., 1996; Kuznetsov, Matrosov, 1997; Кузнецов, Матросов, 2003). При этом создаются условия для поэтапного слежения за переходами эктопаразитов с одного меченого грызуна на другого в случае многократной регистрации конкретных экземпляров блох.
Разнос блох в поселениях различных грызунов-носителей возбудителя чумы активно изучали в 60-70 годы прошлого века. Большинство исследований проведено в Среднеазиатском пустынном очаге чумы, где основным носителем является большая песчанка (Свиридов с соавт., 1961; Руденчик, 1963; Солдаткин, 1963; Солдаткин с соавт., 1966; Корнеев с соавт., 1967; Новокрещенова с соавт., 1967; Руденчик с соавт., 1967; Рапопорт с соавт., 1976 и другие).
Отдельные работы были посвящены территориям, заселенным сурками, сусликами, малыми песчанками, полевками и т. д. (Косминский, Карандина, 1959; 1964; Берендяева с соавт., 1966; Лабунец с соавт., 1966; 1974; Петров с соавт., 1968; Иванов, Мокриевич, 1971; Петров, Гончаров, 1971). В ВолгоУральских песках вопрос о разносе блох полуденными и гребенщиковыми песчанками был разработан явно недостаточно (Мокриевич с соавт., 1969; Медзыховский, Бараева, 1974).Полученные нами результаты испытания метода индивидуального мечения блох и изучения форезии этих паразитов в указанном регионе (Кузнецов с соавт., 1993; Кузнецов, Матросов, Козлова с соавт., 1996; Кузнецов, Матросов, 2003), позволили дать более конкретную характеристику этого процесса. Всю работу по мечению блох проводили параллельно изучению перемещений полуденных и гребенщиковых песчанок на стационарных участках наблюдений в Волго-Уральских песках в весенние и осенние сезоны 1991, 1993 и 1994 гг. За этот период одновременно с мечением грызунов было помечено 1617 блох 6 видов. Массовыми видами были блохи N. Iaeviceps (1324 экз.) и X. Conformis (226 экз.), разнос которых осуществлялся песчанками рода Meriones,а в ряде случаев и серыми хомячками (С. migratorius).
При систематическом отлове на участке песчанок (изредка и других грызунов), их ежедневно приносили в лабораторию непосредственно в живоловках, очесывали с помощью марлевого конуса (Трухачев, Федорова, 1967), а блох с каждого хозяина помещали в отдельные
293 пронумерованные пробирки. Затем блох из пробирки высыпали в глубокую паразитологическую кювету, брали по 1-2 экз. и в отдельной, только для этого используемой пробирке, обездвиживали парами эфира (не более 25-30 сек.). Под микроскопом в чашке Петри с помощью кинжально заточенной препаровальной иглы у них отсекали определенные членики лапок и возвращали в свою пробирку уже с небольшим количеством песка. После полного пробуждения эктопаразитов и не более чем через 30 мин. после мечения последнего, всех животных относили на участок.
Перед выпуском на месте поимки зверька сажали в банку с полиэтиленовой крышкой или пластиковую бутылку, имеющие отверстие под пробирку. Блох, счесанных с грызуна и получивших персональные номера, высыпали на него через отверстие и контролировали их «исчезновение» в шерсти.Индивидуальное мечение блох заключалось в следующем. Каждой блохе присваивали очередной номер, которому соответствовало количество отсекаемых члеников лапок. Членики левой стороны тела нумеровались от 1 до 5, начиная с последнего к ее основанию, правой — от 6 до 9 (правые базальные не использовались для нумерации). Целостность обеих лапок какой-либо пары ног означало цифру 0. Передние лапки означали единицы, средние — десятки, задние — сотни. В случае ампутации члеников правой лапки, левые из этой же пары ног не отсекались, то есть при необходимости получения трехзначного номера с тремя значащими цифрами (до 999), травмированными оказывались только 3 лапки. Как показал опыт, блохи с такими номерами обнаруживались повторно не реже, чем с одно- или двузначными. В качестве примера приведем структуру номера 173: отсутствие одного членика на левой задней лапке (100), двух члеников на правой средней (70) и трех - на левой передней лапке (3). Для самок и самцов каждого вида блох вели отдельную нумерацию. Объем материала и обобщенные данные о смене хозяев блохами приведены в таблице 30.
Таблица 30. Объем материала, полученного методом индивидуального мечения блох в 1991-1994 гг. в Волго-Уральских песках
| Виды блох | Пол блох | Помечено блох | Число повторных встреч блох | Показатель переходов на новых хозяев | |
| всего | со сменой | ||||
| Xenopsylla | самки | 152 | 54 | 14 | 0.259÷0.060 |
| Conformis | самцы | 74 | 49 | 14 | 0.286±0.064 |
| всего | 226 | 103 | 28 | 0.272±0.044 | |
| Nosopsyllus | самки | 900 | 163 | 79 | 0.482±0.039 |
| Iaeviceps | самцы | 424 | 64 | 33 | 0.516±0.062 |
| всего | 1324 | 227 | 112 | 0.493±0.033 | |
| Amphipsylla | самки | 31 | 11 | 1 | — |
| Schelkovnikovi | самцы | 13 | 1 | 1 | — |
| всего | 44 | 12 | 2 | — | |
| Nosopsyllus | самки | 6 | — | — | — |
| mokrzeckyi | самцы | 3 | 1 | 0 | — |
| всего | 9 | 1 | 0 | — | |
| самки | 9 | — | — | — | |
| Прочие виды | самцы | 5 | — | — | — |
| всего | 14 | — | — | — | |
| самки | 1098 | 228 | 94 | 0.412±0.033 | |
| Итого | самцы | 519 | 115 | 48 | 0.417±0.046 |
| всего | 1617 | 343 | 142 | 0.414÷0.027 | |
Анализ обобщенных данных с высокой достоверностью показывает, что блохи N.
Iaevicepsпочти в два раза чаще меняли прокормителей, чем блохи X. Conformis (Р < 0.001). Однако окончательные выводы по этому факту делать преждевременно, поскольку X. Conformisвстречались в достаточном количестве только весной 1993 г., а по результатам мечения N. Iaevicepsниже будет показано, что интенсивность смены хозяев в разные годы варьировала достаточно сильно.В 1991 г. разнос блох изучали на стационарном участке «Зулхаш», расположенном в центральной части Волго-Уральских песков (Атырауская обл., Казахстан). Весной (апрель-май) и осенью (октябрь) было помечено 630 блох A. Iaeviceps(Кузнецов с соавт., 1993; Кузнецов, Матросов, 2003). Из этого количества 41 экз. (6.5%) встречался разное число раз после
295 мечения. По одному разу встречено 35 экз., по два — 5 экз. и по три раза — 1 экз. Общее число зарегистрированных повторов составило 48.
При повторных обнаружениях блох N. Iaevicepsустановлено, что они весьма активно меняли прокормителей и с их помощью перемещались по территории участка на значительные расстояния за довольно короткий срок. Так, из числа всех повторов 38 (79.2%) оказались осуществленными со сменой хозяина, причем такой процент переходов на других зверьков отмечен и весной, и осенью. Интенсивность смены прокормителей блохами мы предлагаем оценивать показателем переходов, который равен отношению числа обнаружений меченых насекомых на новых зверьках к общему числу повторных находок за любой срок наблюдений и при различных интервалах между регистрациями. В приведенном примере показатель переходов равен 0.79 (таблица 31).
Таблица 31. Результаты мечения блох Nosopsyllus Iaevicepsв ВолгоУральских песках весной и осенью 1991 г. на участке «Зулхаш»
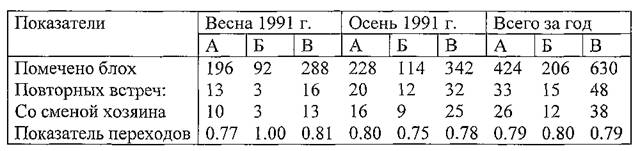
Примечание. А — самки, Б — самцы, В — всего блох.
Анализируя данные за 1991 г., была установлена стабильность параметров смены хозяев блохами N. Iaevicepsи весной, и осенью (около 0.8).
При переходе на другого зверька блохи транспортировались в среднем на 118 м, максимально — на 260 м (поперечник участка) через 7,0 суток (в среднем), при колебаниях от 1 до 19. Таким образом, блохи оставались вне поля нашего зрения довольно продолжительное время, за которое они могли несколько раз поменять хозяев и быть перенесенными на большое расстояние за границы площадки. То, что значительная частьмеченых блох «уходит» за пределы участка подтверждается тем, что повторно удается найти лишь небольшое их число (5-7%). Быстрый «уход» блох с территории мечения в 1991 г. вполне может быть объяснен зарегистрированным в то время высоким показателем смены прокормителей.
Важным фактом является также то, что среднее фиксируемое расстояние переноса блох грызунами в 2-3 раза больше среднего расстояния между пунктами отлова зверьков. Это лишний раз подтверждает возможность более быстрой, но скрытой эстафетной передачи блох между моментами случайного их обнаружения.
Варианты смены хозяев отдельными экземплярами блох с указанием расстояний переноса представлены в таблице 32 (весна 1991 г.) и таблице 33 (осень 1991 г.).
Таблица 32. Характеристика циркуляции блох в поселениях малых песчанок Волго-Уральских песков весной 1991 г.
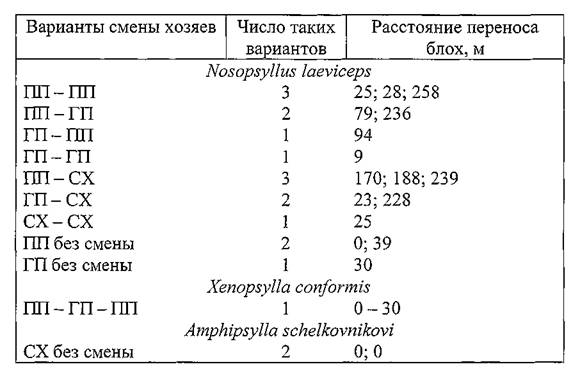
Примечание. ПП — полуденные и ГП — гребенщиковые песчанки; CX — серые хомячки.
Таблица 33. Характеристика циркуляции блох в поселениях малых песчанок Волго-Уральских песков осенью 1991 г.
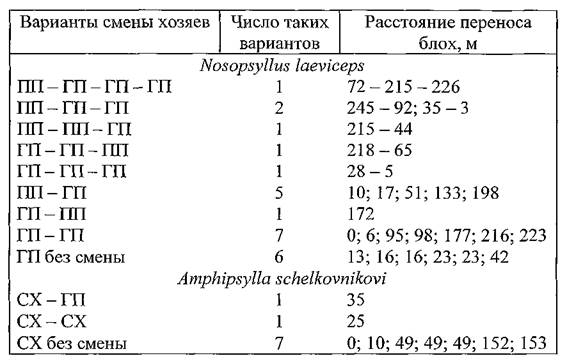
Примечание. ПП — полуденные и ГП — гребенщиковые песчанки; CX — серые хомячки.
Осенью 1993 г. и весной 1994 г. было помечено 682;?-блохи N. Iaeviceps(таблица 34) на участке «Новая Школа» (Кузнецов, Матросов, Козлова с соавт., 1996). После мечения встречено 139 экземпляров (20.4%), что в 3 раза выше аналогичного показателя в 1991 г. Один раз встречено 106 меченых эктопаразитов, два раза — 28, три раза — 3 и четыре раза — 2. В сумме это составило 179 повторов. В 74 случаях зарегистрированы переходы на новых прокормителей, что соответствует показателю, равному 0.41. Здесь отмечена несколько меньшая, чем в 1991 г. стабильность этого показателя в различные сезоны и на другом, более низком уровне. Показатель перехода на нового хозяина самками и самцами блох практически не различался. Однако ив 1991, и в 1993-1994 гг. самки блох преобладали в очесах над самцами в 2 раза, из чего можно заключить, что их приуроченность к телу хозяина также в 2 раза выше, чем самцов (из расчета равновесного соотношения полов). На этом участке при переходе на других зверьков блохи N. Iaevicepsтранспортировались в среднем на
117 м (максимально — на 275). Повторные обнаружения регистрировались в среднем через 5 суток, при колебаниях от 1 до 39. В случае смены хозяина средний интервал между обнаружениями составил 7 суток. Отмечено также повышение доли повторно обнаруживаемых паразитов с параллельным снижением показателя переходов на новых зверьков.
Таблица 34. Результаты мечения блох N Iaevicepsв Волго-Уральских песках осенью 1993 г. и весной 1994 г. на СУН «Новая Школа»
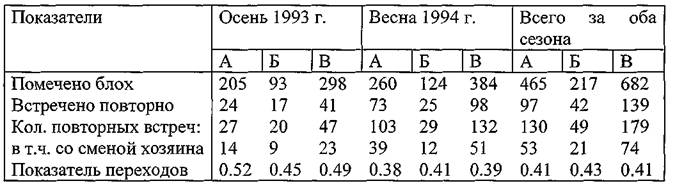
Примечание. А — самки, Б — самцы, В — всего блох.
Весной 1993 г. на СУН «Новая Школа» значительно преобладали блохи X. Conformis,которых помечено 204 экз. (таблица 35). Блох N. Iaevicepsбыло помечено всего лишь 12 экз. и повторно не обнаружено ни одной. В данном случае прослеживается наличие конкуренции между разными видами блох (Чумакова, Ермолова, 2000), которая нейтрализуется их адаптацией к паразитированию в разные сезоны (Агеев с соавт., 1983).
Из числа меченых блох X. Conformis 63 особи (30.9%) встречались и в последствии. Один раз встречено 37 экз., два раза — 18 экз., три раза — 4 и четыре раза — 3 экз. В сумме это составило 99 повторных обнаружений. Из этого числа 26 были со сменой прокормителя, что соответствовало показателю переходов, равному 0.26. Эти находки фиксировались в среднем через 5 суток. Все повторные обнаружения регистрировались в среднем через 2.8 суток при колебаниях от 1 до 13. При переходе на других зверьков блохи X. Conformisтранспортировались в среднем на 94 м (максимально — на 240).
Таблица 35. Результаты мечения блох X. Conformisв ВолгоУральских песках весной 1993 г. на СУН «Новая Школа»

Как уже было отмечено, в различные сезоны наблюдалась разная доля повторно обнаруживаемых блох. При этом интенсивность смены прокормителей блохами также заметно разнилась. Между указанными величинами, которые приведены в таблице 36, обнаружена сильная обратная связь (г = - 0.96), — при значительном уменьшении интенсивности смены хозяев существенно увеличивалось количество повторно обнаруживаемых меченых эктопаразитов. Это происходило независимо от вида насекомого, хотя сама частота переходов блох с одних зверьков на других, очевидно, зависит от свойственного виду режима паразитирования. На наш взгляд, причиной этого является то, что высокая частота смены прокормителей повышает скорость «эмиграции» блох за пределы участка наблюдений, при которой большее их количество успевает «уйти» из-под контроля.
Таблица 36. Зависимость доли обнаруживаемых повторно блох от интенсивности смены ими своих прокормителей 
Вместе с понятием об интенсивности переходов блох на новых хозяев, выражаемой показателем переходов, необходимо ввести термин «вероятность смены прокормителя» и расшифровать его содержание. Временный паразит (блоха) в течение жизни многократно нападает на хозяина и столько же раз покидает его. Если очередное нападение совершается на другую особь относительно предыдущего нападения, то этот факт означает смену прокормителя. Вероятность (частота) смены хозяина равна отношению числа нападений со сменой хозяина к общему их числу, не считая первого нападения, либо за всю жизнь блохи, либо за какой-то ее отрезок. Однако определить полный перечень прокормителей конкретной блохи практически невозможно. Поэтому нужное значение получают, используя данные о двух последовательных (в пределах соседних трофических циклов) нападениях у большого количества блох.
Используя данные о смене хозяев, полученные в природных условиях, важно вычленить из них следующие друг за другом нападения отдельных блох. Единственной возможностью сделать такое вычленение было рассмотрение случаев обнаружения блох с интервалом в одни сутки. Такой срок вероятнее всего соответствует продолжительности одного трофического цикла блохи (Кузнецов, 1997 а) и обеспечивает вычисление искомой вероятности смены прокормителя.
При увеличении интервала между регистрациями случаи перехода эктопаразитов на других зверьков учащаются и уже не могут характеризовать вероятность смены хозяина. За любой срок, превышающий трофический цикл блохи, формируется менее строгий показатель переходов на новых хозяев, который может использоваться для анализа и сравнения различных ситуаций при условии его получения через сопоставимые интервалы времени.
За период наших исследований отмечено 105 случаев двух последовательных находок меченых блох с интервалом в сутки. Из них лишь 13 обнаружений оказались со сменой хозяина, что соответствует
ЗОЇ интегрированной вероятности, равной 0.124. Однако эта величина не так уж мала, поскольку довольно быстро показатель переходов на новых хозяев возрастает и через 10-15 дней приближается к 1.0.
Показатель переходов эктопаразитов на других хозяев зависит от вероятности смены прокормителя при каждом новом нападении и от величины интервала времени между регистрациями. Чем продолжительнее срок между двумя находками блохи, тем больше нападений она может совершить за этот период и тем вероятнее, что во второй раз она будет обнаружена на другом зверьке. Хотя вероятность смены хозяев у изученных видов существенно различалась, через 4 и более суток показатели их переходов практически уравнивались.
Данные о влиянии интервала между регистрациями меченых блох на показатель смены ими хозяев приведены отдельно по видам и участкам в таблице 37 (дробление материала несколько снижало представительность отдельных выборок). Число вариантов с 4-суточным или более длинным интервалом колебалось от 1-2 до 10-15, поэтому для помещения в таблицы они были объединены. По данным таблицы хорошо видно, что при увеличении интервала между регистрациями блох возрастала частота их обнаружения на новых хозяевах. Если блох вновь регистрировали через 15 и более дней, то практически все они были уже на других зверьках.
Анализ отдельных переходов блох с одного грызуна на другого показал, что достаточно часто при этом происходила смена вида прокормителя (таблица 38). Резкое преобладание переходов с участием гребенщиковых песчанок объясняется более высокой численностью этих грызунов на участке мечения в период исследований, особенно на СУН «Новая Школа», и повышенными индексами обилия блох на них. Эти же данные подтверждают отсутствие выраженной специфичности блох N. Iaevicepsи X. Conformisк какому-либо виду малых песчанок и о беспрепятственном «перемешивании» их микропопуляций в поселениях этого двувидового сообщества.
Таблица 37. Вероятность смены прокормителя и показатели переходов блох N. Iaevicepsи X. Conformisчерез разное число суток в Волго-Уральских песках
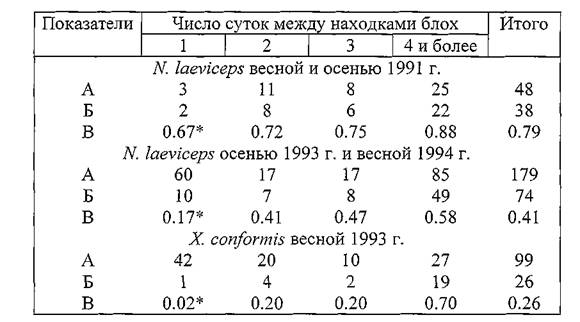
Примечание. А — количество повторных находок блох; Б — в том числе со сменой хозяина; В — вероятность смены хозяина (*) и показатели переходов блох.
Таблица 38. Количество различных вариантов смены прокормителей блохами в поселениях малых песчанок Волго-Уральских песков

Примечание. ПП — полуденные и ГП — гребенщиковые песчанки; CX — серые хомячки.
Показательным явился анализ схем форезии тех экземпляров блох, которые попадались 3-5 раз. Таких насекомых зарегистрировано 67. Не все
эти блохи меняли хозяев или не все повторные нападения отдельных особей сопровождались сменой зверька, поэтому общее число переходов на новых прокормителей в этой группе оказалось равным 58 (по сумме видов N. Iaevicepsи X. conformis).Из особей, попадавшихся 5 раз (5 экз.), интересна судьба самки блохи X. Conformis№ 20 (рисунок 29), которая весной 1993 г. сначала была обнаружена на гребенщиковой песчанке (ГП) № 4 и помечена, а через 5 дней она была счесана с полуденной песчанки (ПП) № 1 в 10 м от места мечения. Еще через ночь — обнаружена на той же ПП № 1 в 20 м от предыдущего места, затем спустя 3 дня на ГП № 2, преодолев 108 м, а еще через 4 дня — снова на IJLll № 1 на прежнем месте, то есть, преодолев те же 108 м в обратном направлении. В приведенном примере зафиксирован случай возврата на одного из прежних хозяев. Подобные возвраты, если их промежуточные этапы по случайным причинам ускользают от регистрации, способны несколько снизить вычисляемый показатель переходов на других прокормителей.
Другой случай возврата на одного из прежних хозяев зафиксирован осенью 1991 г. Самка блохи N. Iaeviceps№ 123 (рисунок 30) сначала была обнаружена на ПП № 116, через 6 дней ее счесали с ГП № 46 в 72 м от места мечения, еще через 3 дня она была встречена на ГП № 62 в 215 м от места предыдущей регистрации, а затем через 1 день — снова на ГП № 46, переместившись почти в обратном направлении еще на 226 м.
Характерно, что маршруты перемещений блох, попадавшихся четыре-пять раз, имели резко изломанную конфигурацию, как, например, на рисунке 31. Очевидно, это обстоятельство способствовало более длительной задержке данных экземпляров на участке и позволило нам их обнаружить несколько раз. Вероятно, более «прямые» маршруты обеспечивали «уход» насекомых с территории участка.
304
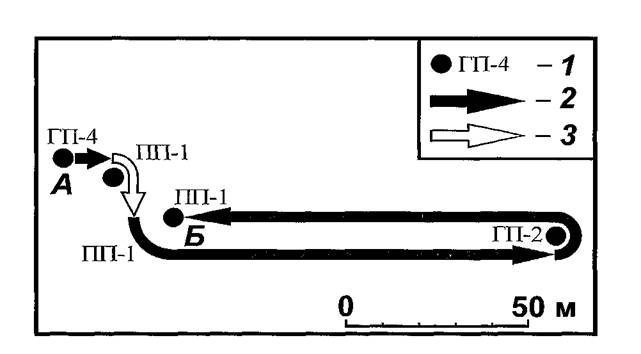
Рис. 29. Схема последовательных обнаружений самки блохи Xenopsylla conformis№ 20 весной 1993 г. на СУН «Новая Школа» в ВолгоУральских песках. 1 — места вылова хозяев, с которых была счесана блоха (А — первое, Б — последнее); перенос блохи со сменой хозяина (2) и без смены (3).
305
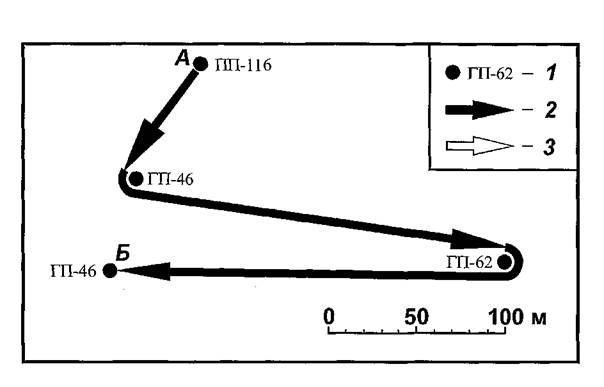
Рис. ЗО. Схема последовательных обнаружений самки блохи Nosopsyllus Iaeviceps№ 123 осенью 1991 г. на СУН «Зулхаш» в ВолгоУральских песках. 1 — места вылова хозяев, с которых была счесана блоха (А — первое, Б — последнее); перенос блохи со сменой хозяина (2) и без смены (3).
306
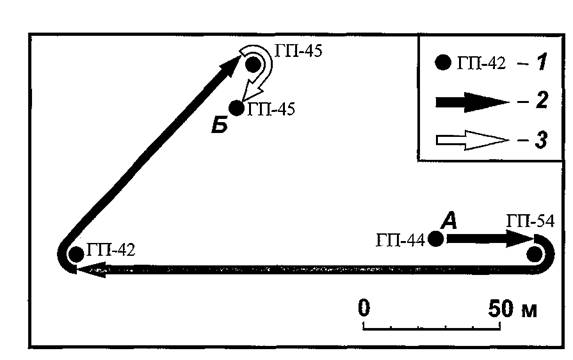
Рис. 31. Схема последовательных обнаружений самки блохи Nosopsyllus Iaeviceps№ 106 весной 1994 г. на СУН «Новая Школа». 1 — места вылова хозяев, с которых была счесана блоха (А — первое, Б — последнее); перенос блохи со сменой хозяина (2) и без смены (3).
Схемы форезии блох позволяют судить о том, какие грызуны на участке осуществляют друг с другом норовые контакты. Нами были отмечены «пары» зверьков (не обязательно одного вида или разного пола) между которыми происходил обмен блохами, причем неоднократный. Экземпляры, помеченные на одном зверьке, затем обнаруживались на другом, а экземпляры, помеченные на втором, приблизительно в эти же дни переходили на первого. При этом не имеет существенного значения, кто транспортирует блох в каждом конкретном случае — предыдущий ли хозяин приносит их в нору нового прокормителя или будущий «подбирает» эктопаразитов в норе предшественника.
Неоднократно были зафиксированы «разветвленные» передачи блох, когда на одном зверьке было помечено несколько особей, а через некоторое время они обнаруживались на разных носителях и в разных частях участка. Отмечены и противоположные факты, когда помеченные в разных местах и на разных грызунах блохи затем были найдены вместе на другом животном («сходящиеся» передачи).
Обнаружены факты и иного рода. Когда блохи, помеченные на одном грызуне, через некоторое время вместе друг с другом оказывались на новом зверьке, то есть имели как бы «сцепленную» судьбу. Очевидно, покинув прокормителя в одной норе, они напали на нового одновременно, что говорит о синхронизации трофических циклов и привязке их к определенным моментам суточной активности грызунов. Отмечены также факты, когда одна из нескольких помеченных одновременно на конкретном зверьке блох через некоторое время регистрировалась на другом хозяине, а еще через один или несколько дней на этом же новом хозяине обнаруживалась другая блоха из той же первоначальной группы меченых. Это говорит о существовании некоторой «стандартности» в норовых контактах грызунов и служит подтверждением консерватизма песчанок не только в суточной активности, но и в ежедневно пробегаемых маршрутах.
Простой и наглядной иллюстрацией перечисленных вариантов паразитарных связей может служить схема передач 9 экземпляров блох N. Iaeviceps,осуществленных в мае 1991 г. при помощи 8 особей грызунов: трех полуденных и двух гребенщиковых песчанок, а также трех серых хомячков. В этот сезон 13 блох меняли прокормителей, но 9 из них вошли в единую «сеть» передач (рисунок 32). Две «стандартизованные» передачи блох № 15 и № 9, помеченных 9 мая 1991 г. на Illl № 69, осуществились на CX № 4 сначала 24 мая (блоха № 15), затем 28 мая (блоха № 9). Из трех «расходящихся» передач одна случилась в той же группе блох, меченных 9 мая на ПП № 69. Блоха № 9 перешла на CX № 4, а блоха №7 — на ГП № 10. Другая пара «разошедшихся» блох была помечена 18 мая на ГП № 41. Блоха № 48 перешла затем на ГП № 10, а блоха № 50 — на CX № 4. Третья пара блох была помечена 9 мая на ПИ № 61, одна из которых затем была обнаружена 18 мая на ПП № 71, другая — 22 мая на CX № 3. «Сходящихся» пар блох зарегистрировано также три. Блохи № 9 и № 50 встретились 28 мая на CX № 4, хотя одна из них (№9) была помечена на ПП № 69, другая — на ГП №41.
Другие две блохи (№ 7 и № 48) помечены на тех же песчанках, хотя затем встретились 20 мая на ГП № 10. Третья пара блох (№ 60 и № 90) «сошлись» на CX № 5, хотя за три дня до этого были помечены на ГП № 41 иСХ№3.
Выявление столь разветвленной и «тесной» связи между грызунами и блохами выглядит «как чудо» и обязывает сделать из этого определенные выводы. Прежде всего, отметим, что поимки меченых грызунов, а тем более блох — события исключительно случайные и редкие. Несмотря на это выявленные паразитарные контакты в короткий срок охватили значительную часть участка наблюдений.
309
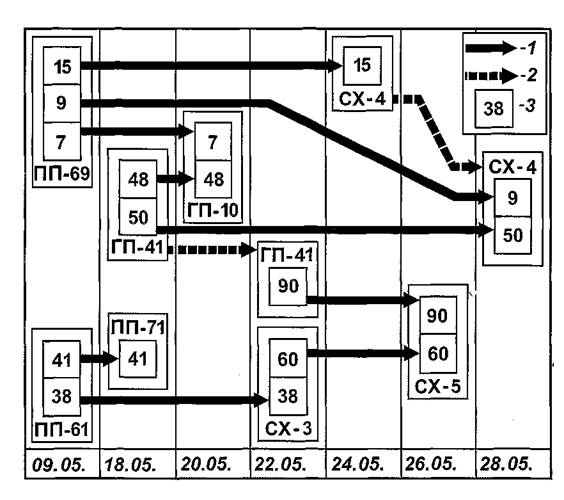
Рис. 32. Схема «контактной сети» в поселениях малых песчанок, серых хомячков и блох Nosopsyllus Iaevicepsвесной 1991 г. на СУН «Зулхаш». 1 — путь смены хозяина блохой; 2 — путь, связывающий участие зверька в передаче разных блох в разное время; 3 — номера блох. Номера и аббревиатуры прокормителей — см. в тексте.
В реальных условиях передвижения и норовые контакты грызунов происходят ежесуточно. Расстояния в 100, 200 и более метров, как мы уже выяснили, не являются препятствием для этого. Поэтому истинные масштабы контактов во столько же раз более объемны, во сколько раз выборка, составленная из моментов регистрации меченых животных, меньше генеральной совокупности событий в их реальной жизни. Все это говорит о том, что группировки грызунов — обитателей открытых ландшафтов и популяции их эктопаразитов весьма обширны в пространственном отношении и монолитны (непрерывны) в «социальном» смысле.
В другие периоды исследований были зарегистрированы уже известные варианты передач, обеспечивающие формирование «контактной сети». В октябре 1991 г. были выявлены три фрагмента сети, формально не связанные между собой, что следует расценивать не как фактическое отсутствие связи, а как недобор данных. Наибольший фрагмент был образован переходами 10 экз. блох N. Iaeviceps,«связавших» между собой 13 особей песчанок двух видов (рисунок 33). Осенью, так же как и весной, наблюдались «сходящиеся» и «расходящиеся» передачи.
Еще более эффектные результаты получены весной 1993 г. на участке «Новая Школа» при мечении блох X. conformis. В смене прокормителей участвовали 22 блохи, осуществившие в общей сложности 26 переходов. Оказалось, что все эти эктопаразиты вошли в единую сеть паразитарных контактов, представленную всеми формами переходов. Передачи и распространение блох по участку осуществляли 1 полуденная и 6 гребенщиковых песчанок. Замечены неоднократные «сходящиеся», «расходящиеся» и повторяющиеся («стандартные») передачи, «сцепленные» передачи и сложные комбинации этих форм у отдельных экземпляров блох. Все это происходило при относительно низкой численности песчанок, однако индексы обилия блох были достаточно высокими. Среднее расстояние переноса блох в случае смены хозяина
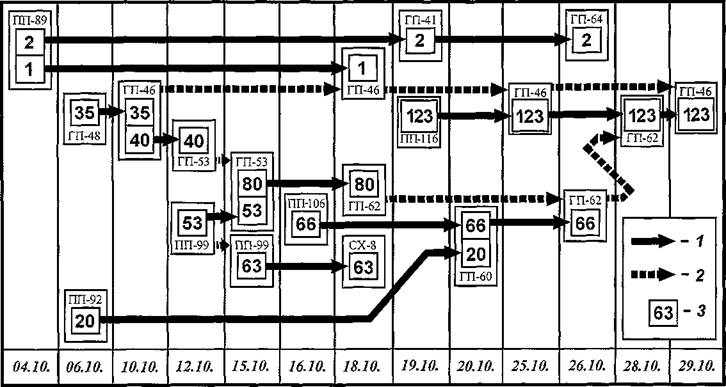
Рис. 33. Схема «контактной сети» в поселении полуденных и гребенщиковых песчанок и блох Nosopsyllus Iaevicepsосенью 1991 г. на СУН «Зулхаш». 1 — путь смены хозяина блохой; 2 — путь, связывающий участие зверька в передаче разных блох в разное время; 3 — номера блох. Номера и аббревиатуры прокормителей — см. в тексте.
312 составило 94 м, что несколько меньше, чем в другие сезоны, но позволило связать воедино норы на всем участке.
Таким образом, всего лишь за 16 дней работы весной 1993 г. на участке, площадью 9 га, где обитало не более 9 малых песчанок и помечено 204 экз. блох X. conformis,был выявлен полный охват этой территории паразитарными связями между всеми грызунами и паразитирующими на них блохами. Поскольку поселения песчанок продолжались и за пределами участка без ощутимых разрывов, можно предположить, что непрерывная сеть контактов простиралась во все стороны, повторяя расположение нор грызунов. Заметим, что этот вывод подтверждался и в другие периоды исследований на обоих участках.
Осенью 1993 г. на том же участке меняли прокормителей 21 блоха N. Iaeviceps.Все они вошли в единую контактную сеть, в формировании которой приняли участие 16 гребенщиковых песчанок из 30 особей, обитавших на участке. Здесь также отмечены все варианты переходов, охватившие всю площадь участка наблюдений. В среднем блохи переносились на 134 м.
Наиболее обширная, разветвленная и монолитная сеть паразитарных контактов выявлена весной 1994 г., благодаря относительно
продолжительному периоду работы (с 10 апреля по 31 мая), в которой блохи, менявшие прокормителей, регистрировались в течение 25 дней. В обнаруженных контактах были задействованы 19 гребенщиковых песчанок и 47 блох N Iaeviceps.
Весной и осенью 1993 г., а особенно весной 1994 г. образовавшиеся сети контактирования оказались настолько сложными и обширными, что было решено не приводить в диссертации трудные для вычерчивания и восприятия схемы. Скажем лишь, что во всех этих сетях были многократно представлены все виды передач.
Таким образом, при помощи метода индивидуального мечения блох удалось предметно и объективно охарактеризовать уровень
313 .
внутривидовых и межвидовых паразитарных связей грызунов и блох. Материалы исследования говорят о том, что паразитарные контакты между полуденными и гребенщиковыми песчанками, то есть между особями как одного, так и разных видов, осуществляются ежесуточно и беспрепятственно. При этом происходит закономерный переход блох C одних зверьков на других и их перенос на 100, 200 и более метров. Высокий уровень контактирования отмечался во все сезоны, несмотря на то, что численность песчанок и их блох заметно менялась. Несомненно, что поддержание внутрипопуляционных контактов на оптимальном уровне является насущной (жизненной) потребностью животных. Контакты, по всей вероятности, осуществляются всегда и везде, где отсутствуют разрывы в поселениях, превышающие «мобильные» возможности грызунов.
Все приведенные выше факты свидетельствуют о высокой степени «обобществления» эктопаразитов в пределах, как минимум, каждого цельного поселения грызунов, а, возможно, и на более обширных пространствах, адекватных ареалам популяций прокормителей. Причем в Волго-Уральских песках функцию «несущей» популяции выполняют совместные поселения полуденных и гребенщиковых песчанок, представляющие единое целое для блох, паразитирующих на хозяевах этих поселений. Наши материалы служат убедительным подтверждением справедливости такого же мнения, высказанного А.С. Бурделовым с соавт. (1985).
Насекомые, которые не успели сменить прокормителей, транспортировались песчанками в среднем на меньшие расстояния. Это вполне объяснимо, поскольку зверек двигается в пределах своего индивидуального участка и пока блоха находится на нем, дистанции их перемещений совпадают. Так, по данным за все сезоны средний перенос блох со сменой хозяина составил 111м, без смены — 49 м. Последнее значение соответствует среднему радиусу ежесуточных перемещений
песчанок. Однако при вычленении из данных о переносах блох со сменой хозяина случаев, зарегистрированных через 1 сутки (13 измерений), среднее расстояние переноса блох оказывается даже большим и составляет 126 м при колебаниях от 15 до 242 м. Регистрация, сопровождавшаяся переходом на нового прокормителя, наблюдалась в среднем почти через 7 суток, без перехода - через 3 суток. Кроме того, были отмечены особенно большие сроки между регистрациями блох, сопровождавшимися сменой хозяина, вплоть до 22-39 дней. При этом маловероятно, что паразиты в эти дни не питались, поскольку им свойственны практически ежедневные кровососания. Одной из возможных причин их долгого «необнаружения» могла быть «эмиграция» за пределы участка, а следовавшие за этим возвращения фиксировались нами через 3-5 недель.
Таким образом, в результате проведенных нами исследований в Волго-Уральском песчаном очаге чумы мы обнаружили, что разнос блох
N. Iaevicepsи X. conformisосуществляется обитающими там полуденными и гребенщиковыми песчанками в очень широких масштабах. В среднем через неделю около половины эктопаразитов оказываются на других прокормителях, преодолевая с их помощью более 100 м. Усредненная вероятность смены хозяина блохами в течение суток (что адекватно одному трофическому циклу) оценивается как 0.124, однако, в отдельные сезоны у разных видов блох она может меняться в широких пределах (от
O. 02 до 0.81).
5.2.3.
Еще по теме Форезия блох в Волго-Уральских песках:
- Перемещения полуденных и гребенщиковых песчанок в Волго-Уральских песках.
- Волго-Уральский песчаный очаг(16)
- Форезия блох в сельских населенных пунктах на плато Тайнгуен
- Структура Волго-Уральского песчаного очага чумы.
- Ландшафтная приуроченность элементов пространственной структуры популяций песчанок в ВолгоУральском песчаном очаге чумы.
- Питание блох.
- Культивирование блох.
- Методы сбора блох.
- Продолжительность метаморфоза блох в днях
- Значение блох в распространении инфекционных заболеваний
- Число блох, питающихся на одном прокормителе в период бактериемии
- РАСПРОСТРАНЕНИЕ ВОШЕДШИХ В ОПРЕДЕЛИТЕЛЬ РОДОВ БЛОХ В СССР И ОСНОВНЫЕ ХОЗЯЕВА г (И. ИОФФ).
- Параграф десятый. Как прогнать блох