1.3 Современные подходы к видовой идентификации возбудителей инвазивного кандидоза
Подтверждение диагноза ИК в России базируется на следующих критериях диагностики: получение роста Candida spp. при посеве крови и других в норме стерильных биосубстратов, биопсийного материала от пациента с клиническими признаками инфекции, а также при обнаружении Candida spp.
при гистологическом исследовании глубоких тканей. Для получения культуры возбудителя ИК забор крови производят у пациентов с температурой более 380С или другими признаками генерализованной вопалительной реакции 2 раза в день в течение не менее 3 дней, серологические тесты не рекомендованы. Посев производят на жидкие питательные среды (бульон Сабуро, сусло) [4]. В соответствии с Европейскими рекомендациями по диагностике кандидоза (2012г.), заборкрови производится ежедневно трехкратно из каждой локтевой вены с интервалом 30 минут, посев осуществляют во флаконы систем BacT/ALERT и других (для аэробных и анаэробных культур) с инкубацией не менее 5 суток [63], что позволяет определить наличие или отсутствие Candida spp. в крови [37].
Другие стерильные в норме биоматериалы засевают также на плотные питательные среды. Для этого в большинстве лабораторий используют агар Сабуро, сусло – агар. Применение хромогенных сред – питательных сред с субстратом, который, взаимодействуя с ферментами различных видов Candida, окрашивают их колонии в определенные цвета (CHROMAgar
Candida, BD Diagnostic) [136], помогает выявить разные виды Candida в смешанной культуре, однако точно идентифицировать вид Candida не позволяет [5, 21].
При выделении культуры микромицета для видовой идентификации в настоящее время в практике используют различные коммерческие методы, например тест-системы Remel BactiCard Candida Kit (Thermo Scientific™, США). При чувствительности и специфичности данной тест-системы –
98,7 % и 99,6% соответственно, за 5 минут данный тест позволяет дифференцировать вид C.
albicans от других видов Candida [54]. Тест на образование «ростковых» трубок также позволяет отличить C. albicans, C. dubliniensis, образующих ростковые трубки в сыворотке крови, от других видов.Таким образом, заметим, что данных методов идентификации вида недостаточно, поскольку они расчитаны в основном на дифференцировку C. albicans от C. не-albicans видов. Более предпочтительны методы видовой идентификации, ориентированные на определение биохимических свойств, молекулярно-генетических особенностей и белкового масс-спектра профиля микромицета (рисунок 3).
Второе поколение
Третье поколение
Биохимическое направление (автоматические анализаторы )
Молекулярно- генетическое направление (ДНК-секвенирование)
Идентификация грибов
Первое поколение
Четвертое поколение
Биохимическое направление
(ручные тест-системы)
Физико-химическое направление (MALDI-TOF масс- спектрометрия)
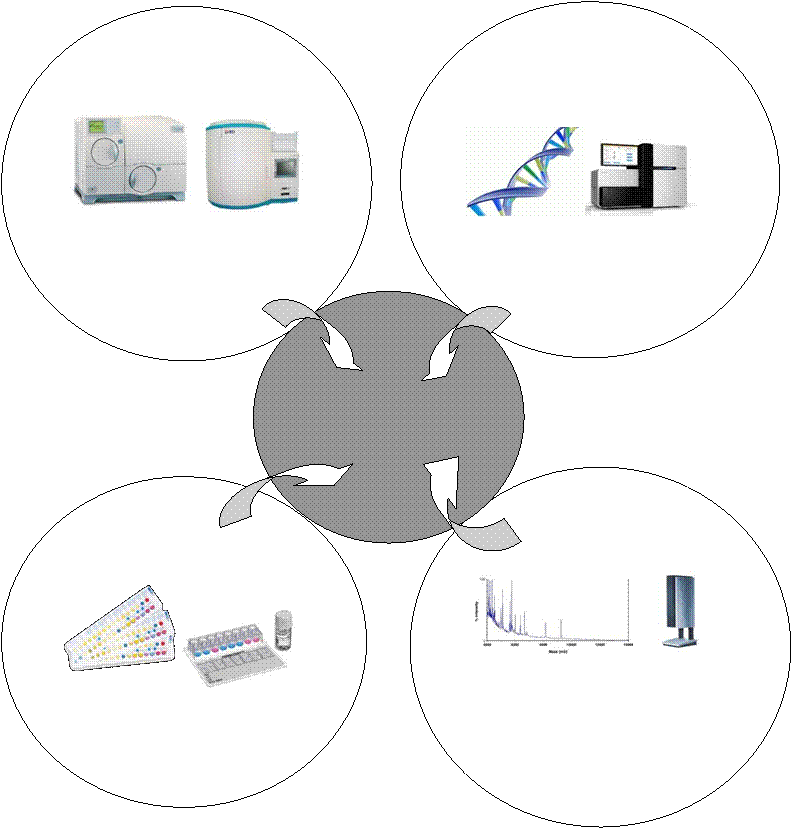 Рисунок 3 – Развитие методов видовой идентификации грибов рода Candida spp.
Рисунок 3 – Развитие методов видовой идентификации грибов рода Candida spp.
Методы видовой идентификации, основанные на изучении молекулярно-генетических особенностей, стали применяться сравнительно недавно, а протеомные технологии – начиная с 2000-х годов [60].
Одним из давно разработанных методов идентификации является определение видов Candida по биохимическим свойствам микромицета. Для определения ферментации углеводов раньше использовали жидкие питательные среды с углеводами, реактивом Андреде и стеклянными поплавками. Ассимиляцию углеводов проводили методом дисков, т.е.
использовали бумажные диски с нанесенными на них растворами углеводов [14]. За последние десятилетия появились промышленно приготовленные тест – системы как для ручной постановки теста, так и для автоматической. К числу «ручных» тест-систем относятся RapID Yeast Plus (Remel), API ID32C, AUXACOLOR2 (BioRad) и другие. Подобные тест-системы просты в использовании и обладают высоким процентом сопоставимости результатов с референтными молекулярно-биологическими методами.
Так, в сравнительном исследовании трех биохимических тест-систем (Candifast, API 20C AUX, Fungichrom) с использованием 116 клинических изолятов, процент правильной идентификации составил 82,7 – 95,6, время определения видов составило от 24 до 48 часов [81].Среди автоматических анализаторов наиболее часто используют Vitek2
Compact (bioMerieux) с картами VITEK2 YST, с их помощью в исследовании проведенном в Италии верно идентифицировали 98,2% из 750 клинических изолятов дрожжей, предварительно прошедших ДНК-секвенирование [76]. Также автоматическими анализаторами являются MicroScan plus WalkAway (Siemens), Phoenix (Becton Dickinson). Выявлено, что совпадение между результатами идентификации MicroScan plus WalkAway и VITEK2 YST составляет 94% [140]. При простой пробоподготовке и высокой сопоставимости результатов с результатами референтного метода (ДНК- секвенирование), длительность инкубации составляет до 18 часов, для анализатора MicroScan plus WalkAway – 4 часа.
Во многих странах проводили сравнительные исследования эффективности биохимических методов идентификации. Так, сравнение тест- систем API ID32C, AUXACOLOR2 и Vitek2 Compact в рамках исследования в отделениях реанимации и интенсивной терапии в Греции в период с 2005 по 2010 гг. выявило, что из 253 изолятов с помощью Vitek2 Compact верно идентифицировано 84%, API ID32C – 83%, AUXACOLOR2 – 80%. Так, среди наиболее часто встречающихся видов, таких как C. albicans, C. glabrata, C. parapsilosis процент верно идентифицированных штаммов был выше (AUXACOLOR2 – 94,5%, API ID32C – 94%, Vitek2 Compact – 91%), чем в
отношении редких видов (AUXACOLOR2 -74%, API ID32C – 75%, Vitek2
Compact – 82%) [87]. С помощью трех систем точно идентифицировали более
91% часто встречающихся изолятов, и эти системы могут быть применены в рутинной практике; для редких видов рекомендовано применение молекулярно-генетических методов. В исследовании, проведенном в Бразилии, при сравнении результатов Vitek2 Compact и API ID20C AUX было обнаружено, что совпадение составляет 93,6 % [158].
Таким образом, основным недостатком методов видовой идентификации возбудителей ВБИК по биохимическим свойствам является длительность идентификации (18 – 48 часов), а также невысокий процент правильной идентификации редких видов.
Молекулярно-генетические методы направлены на определение количественных и качественных характеристик первичной структуры ДНК. В настоящее время разработаны и широко применяются различные методы молекулярной генетики общим числом около ста. Основу подавляющего большинства таких методов составляет полимеразная цепная реакиця (ПЦР). Перечень наиболее часто используемых методов приведен в таблице 1[1].
Таблица 1 – Наиболее распространенные молекулярно-генетические методы
| № п/п | Молекулярно-генетические методы |
| 1 | Полиморфизм длин амплифицированных фрагментов (AFLP) |
| 2 | Полиморфизм длин рестрикционных фрагментов (RFPL) |
| 3 | Расщепление резолвазой (EMD) |
| 4 | Методы, основанные на лигазной реакции (LDL, LCR, Radlock) |
| 5 | Инвазивное расщепление олигонуклеотидов (расщепление Clevase I) |
| 6 | Случайная амплификация полиморфной ДНК (RAPD, AP-PCR) |
| 7 | ПЦР с прямой терминацией синтеза (DT-PCR) |
| 8 | Анализ конформации одноцепочечных фрагментов ДНК (SSCP) |
| 9 | Гетеродуплексный анализ |
| 10 | Секвенирование ДНК |
| 11 | Минисеквенирование |
| 12 | Аллель-специфическая ПЦР |
| 13 | Масс-спектрометрия |
| 14 | ПЦР в реальном времени |
| 15 | Микрочипы |
В зависимости от целей исследования все молекулярно-генетические методы можно подразделить на две группы: методы, направленные на определение структуры ДНК (видовая идентификация, поиск новых мутаций), и на анализ известных генетических последовательностей (полиморфных вариантов генов или идентификация микроорганизмов по референтной последовательности ДНК).
Исторически первым методом идентификации генетических последовательностей стал метод блот-гибридизации (Саузерн-блот), предложенный еще в 1975 году английским ученым Эдвардом Саузерном. С открытием термостабильной полимеразы, в практику молекулярных генетиков вошел метод ПЦР, произведший переворот в генетических исследованиях. Метод основан на многократном избирательном копировании определенного участка нуклеиновой кислоты ДНК с помощью ферментов in vitro. Копирование происходит только того участка, что удовлетворяет заданным условиям и если он присутствует в исследуемом образце. В отличие от амплификации ДНК в живых организмах (репликация), с помощью ПЦР амплифицируются относительно короткие участки ДНК. Метод ПЦР лежит в основе всех методов, применяемых для диагностики и идентификации микромицетов в клинической практике [1].
В настоящее время референтным методом определения видовой принадлежности микромицетов является протокол M18-А Института клинических и лабораторных стандартов (CLSI), по которому идентификацию грибов необходимо проводить с применением ПЦР, с последующим секвенированием генов мишеней, которые имеют достаточно разнородный нуклеотидный состав, с высокой межвидовой дивергенцией. Идентификация устанавливается путем выравнивания анализируемой нуклеотидной последовательности с последовательностями, представлен- ными в GenBank или в других базах данных генетической информации. Общие положения данного подхода для молекулярной биологии предложены Д. Сэмбруком и Д. Расселом в 2001 году (Sambrook & Russell) [141].
GenBank является базой данных всех аннотированных нуклеотидных (ДНК и РНК) и аминокислотных последовательностей, которые они кодируют, представленных в свободном доступе для исследователей всего мира. Эту базу данных поддерживает Американская Национальная библиотека медицины Национального центра биотехнологической информации (NCBI). GenBank коллаборирует с другими информационными банками, такими как Европейская молекулярно-биологическая лаборатория (EMBL) и Банк данных ДНК Японии (DDBJ), осуществляют регулярный обмен данными между этими тремя архивами аннотированных нуклеотидных последовательностей [141].
Для видовой идентификации микромицетов выбран комплекс рибосомальных генов (рисунок 4), содержащих высоко консервативные домены, перемежающиеся с более вариабельными областями, которые содержат видоспецифические «подписи». Для упрощения субъединицы этого комплекса называют рДНК, подразумевая, что это гены, кодирующие рРНК
молекулы.
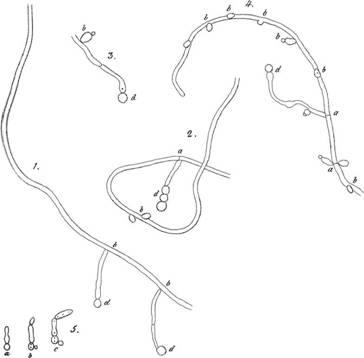 5’ Малая субъединица
5’ Малая субъединица
18S РНК
5.8S РНК
ITS1 ITS2
Большая субъединица
25-28S РНК
IGS1
5S РНК
3’
IGS2
Рисунок 4 – Организация рРНК генов микромицетов
Внутри рДНК комплекса существует совокупность множества тандемных повторов нуклеотидов, находящихся по направлению от 5` к 3` концу между субъединицами 18S и 5.8S, а также 5.8S и 28S, называющихся вариабельными внутренними транскрибируемыми спейсерами (ITS), соответственно 1 и 2. Фланкируют ген большой рибосомальной субъединицы
28S два вариабельных нетранскрибируемых спейсер региона: IGS1-5S-IGS2. У Saccharomyces cerevisiae существует приблизительно 140 единиц повторов. У предшественника транскрипта РНК происходит ферментативное удаление ITS регионов, и, после дальнейших модификаций, формируются 18S, 5.8S и
28S зрелые рРНК, которые в комбинации с белками формируют
функциональные рибосомы. Биологическая роль ITS регионов - участие в ранней транскрипции РНК процессинга. Таким образом, они являются некодирующими регионами генома [141].
ITS1 и ITS2 регионы, фланкирующие 5.8S рДНК ген, показывают значимую нуклеотидную дивергенцию. Каждый ITS регион около 300 пар нуклеотидов в длину. Они очень хорошо подходят для видовой идентификации как дрожжей, так и нитчатых грибов (таблица 2) [90]. Тринадцать известных патогенов, включая потенциально инвазивные виды Aspergillus, можно идентифицировать путем сравнительного анализа их ITS1 и ITS2 последовательностей. При использовании двух спейсерных регионов достигается лучшая межвидовая дифференциация, чем при использовании одного региона.
Таблица 2 – Видовая идентификация путем прямого анализа региона
внутреннего транскрибируемого спейсера (ITS) рДНК [90].
| ITS1 | Wangiella (Exophiala) dermatitidis |
| Epidermophyton floccosum | |
| Microsoporum (4 вида) | |
| Trichophyton (6 видов) | |
| Malassezia (6 видов) | |
| ITS2 | C. albicans, C. parapsilosis, 13 других видов Candida |
| Cryptococcus (5 видов) | |
| Rhodotorula | |
| ITS1 и ITS2 | Aspergillus (13 видов) (Hinrikson et al., 2005) |
| Cryptococcus neoformans | |
| Fonsecaea compacta, F.pedrosol | |
| Histoplasma capsulatum | |
| Paracoccidioides brasiliensis | |
| Pseudallescheria boydii |
Также для видовой идентификации дрожжей используют вариабельный регион (первые, приблизительно 600 пар нуклеотидов) большой
субъединицы РНК 28S (иногда в литературе описывается, как 26S), названный D1/D2 субрегионом, тогда как остальная последовательность 28S высоко консервативна [95]. Последовательности D1/D2 всех видов дрожжей аскомицетов и базидиомицетов известны и представлены в GenBank. Большинство видов дрожжей может быть идентифицировано по их различиям в нуклеотидной последовательности D1/D2 домена. Для восьми видов Candida spp. разработан метод быстрой диагностики с использованием микросфер, с нанесенными нуклеотидными последовательностями D1/D2, для проточной цитометрии [145].
Метод ДНК-секвенирования не зря признан «золотым стандартом» для видовой идентификации микромицетов. По современным данным, видовая идентификация по ITS региону клинических изолятов дрожжей (включая виды Candida) в 99,7% случаев корректна, даже для редких видов [35]. По нашим данным, согласие между результатами молекулярно-генетической и классической фенотипической идентификаций варьирует между исследованиями довольно значительно, в пользу ДНК секвенирования [4].
В настоящее время ДНК-секвенирование используют не только для определения вида внутри рода, но и для типирования внутри вида. Подобное внутривидовое типирование позволяет осуществить подход мультилокусного сиквенс-типирования (MLST), который позволяет изучать, как историческое развитие геномной последовательности, так и приобретенные наследственные характеристики (различные перестройки генома, приводящие к различным фенотипическим вариантам как приобретение резистентных свойств к антимикотической терапии). В исследовании Якобсена и соавторов [80] был проведен анализ изолятов C. albicans MLST, охватывающий семь регионов различных генов. Показано, что между различными сиквенс-типами существует значимая корреляция с географическим нахождением. Данные исследования позволяют определять схемы миграции клинически важных штаммов, и предсказывать эпидемиологические вспышки.
Одним из новых быстрых методов видовой идентификации грибов является технология MALDI-TOF масс-спектрометрии (матрично- ассоциированная лазерная десорбция/ ионизация время-пролетная масс- спектрометрия, от англ. MALDI-TOF MS – Matrix Assisted Laser Desorption/Ionization – Time-of-Flight Mass-spectrometry) – метод видовой идентификации микроорганизмов, основанный на анализе их белкового масс- спектро-профиля [132]. Технология MALDI-TOF масс-спектрометрии была разработана еще в середине 80-х годов учеными К. Танака, Ф. Гилленкампом, М. Карасом и использовалась для изучения строения различных высокомолекулярных соединений, в том числе биологического происхождения [102]. Лишь в начале XXI века метод впервые был применен для видовой идентификации патогенных грибов [101], несколько позже для Candida spp. [102].
В 2008 – 2009 гг. в Германии было проведено сравнительное исследование результатов видовой идентификации Candida spp., полученных методами AUXACOLOR2 и MALDI-TOF масс-спектрометрии, в сравнении с референтным молекулярно-генетическим методом. Обнаружено, что методом MALDI-TOF масс – спектрометрии правильно было идентифицировано 97,6%, а методом AUXACOLOR2 – 96,9% [138]. При этом AUXACOLOR2 не смог выявить виды, входящие в комплекс C. parapsilosis, а изолят C. bracarensis был определен как C. glabrata.
В 2012 году в Италии было проведено исследование по оценке возможности с помощью метода MALDI-TOF масс-спектрометрии определять Candida spp. непосредственно в положительных посевах образцов крови (BC Bactec Mycosis, Becton Dickinson, BD). Полученные данные сопоставляли с результатами видовой идентификации культур Candida из крови, выделенных на плотных питательных средах. Обнаружено, что C. albicans в посевах крови непосредственно в жидкой среде определена в
95,9% образцов, C. не-albicans виды – в 86,5%. Среднее время проведения
анализа составило 30 минут при низких затратах на расходные материалы
[56].
Следовательно, MALDI-TOF масс-спектрометрия является методом быстрой и точной идентификации с низкой себестоимостью. Однако, возможны затруднения в идентификации видов, если их масс-спектры отсутствуют или единичные в базе данных масс-спектрометра.
Чувствительность микромицета к противогрибковым препаратам тесно связана с его видовой принадлежностью, в связи с чем необходим постоянный мониторинг и видового спектра и чувствительности к противогрибковым препаратам, в том числе нового поколения.
Еще по теме 1.3 Современные подходы к видовой идентификации возбудителей инвазивного кандидоза:
- 2.5 Видовая идентификация возбудителей внутрибольничного инвазивного кандидоза с помощью тест-системы AUXACOLOR2
- ГЛАВА 1 Проблемы современной лабораторной диагностики возбудителей инвазивного кандидоза (ОБЗОР ЛИТЕРАТУРЫ)
- 3.6 Морфологические особенности возбудителей инвазивного кандидоза
- 3.1 Источники выделения возбудителей инвазивного кандидоза
- 1.4 Чувствительность возбудителей инвазивного кандидоза к противогрибковым препаратам
- 3.8 Молекулярно-генетические особенности возбудителей инвазивного кандидоза
- 1.2 Грибы рода Candida - возбудители инвазивного кандидоза
- 3.7 Протеомное профилирование возбудителей инвазивного кандидоза
- 2.6 Изучение морфологических особенностей возбудителей инвазивного кандидоза
- 4.1 Результаты определения чувствительности возбудителей инвазивного кандидоза методом CLSI M27-A3.
- 4.2 Результаты определения чувствительности возбудителей инвазивного кандидоза методом CLSI M44 - A
- 4.3 Результаты определения чувствительности возбудителей инвазивного кандидоза с помощью анализатора Vitek2 Compact
- Видовой состав возбудителей Malassezia-инфекций и их распространенность у животных различных видов
- 3.2.2 Видовая идентификация Cryptococcus neoformans
- Видовая идентификация и биологические особенности грибов рода Malassezia
- 3.2 Сравнение результатов видовой идентификации Candida spp. методами ДНК-секвенирования и MALDI-TOF масс-спектрометрии
- Современные представления о вагинальном кандидозе
- 3.4 Видовой спектр возбудителей ВБИК в течение 2012-2014гг.
- 4.1 Видовой состав оводов - возбудителей гастерофилеза лошадей