Влияние ацилированных производных дофамина и серотонина на гидролиз анандамида
Известным способом регуляции действия нейролипинов из группы этаноламидов жирных кислот (например анандамида) является их деградация ГАЖК, экспрессируемой в тканях головного мозга (Deutsch et al., 2002).
Устойчивость производных жирных кислот с дофамином и серотонином, а также взаимодействие этих соединений с ГАЖК практически не изучались.Методом тонкослойной хроматографии нами было показано, что при инкубации анандамида с препаратами мембран головного мозга крысы в органических экстрактах смеси происходит повышение содержания свободной арахидоновой кислоты, что свидетельствует о гидролизе данных соединений. Использование ингибитора сериновых амидгидролаз фенилметилсульфонилфторида приводило к значительному снижению образования арахидоновой кислоты из анандамида. При инкубации АДА в тех же условиях обнаруживалось незначительное, по сравнению с анандамидом, количество арахидоновой кислоты. N-арахидоноилсеротонин (АСТ) был полностью устойчив к гидролизу. Анализ экстрактов с помощью ВЭЖХ также подтвердил стабильность АСТ в данных условиях. Был проведен анализ влияния АДА и АСТ, а также их производных на активность ГАЖК в препаратах мембран головного мозга крысы (Bisogno et al., 1998, 2000; Бобров и др., 2000; Безуглов и др., 2006; Генрихе и др., 2010). В качестве субстрата использовали [3Н]-анандамид, меченный по остатку этаноламина (Рогов и др., 1997). Среди N-ацилдофаминов наибольшую ингибиторную активность проявлял N-эйкозапентаеноилдофамин (1с-4Ь), тогда как АДА был наименее активным. Таким образом, в этом тесте производные ю3 жирных кислот были активнее производных ю6 кислот (табл. 2).
Среди N-ацилсеротонинов наибольшей активностью обладали N-эйкозапетае- ноил-серотонин (ЭСТ, 1с-4Ь) и АСТ (1d-4b) с IC50 1 мкМ и 5,2 мкМ, соответственно (табл. 3). По-видимому, молекулы, содержащие эйкозапентаеновую кислоту, имеют наиболее подходящую конформацию для взаимодействия с ферментом.
Таким образом, замена арахидоновой кислоты в составе нейролипинов на ю3 жирные кислоты может приводить к усилению ингибирования ГАЖК.Можно предположить, что остатки дофамина и серотонина в молекулах ней- ролипинов обеспечивают ингибирующее действие за счет гидроксильных групп
Таблица 2
Ингибирование ГАЖК N-ацил-дофаминами и их аналогами. Сопоставление с активностью этих соединений в отношении КР1
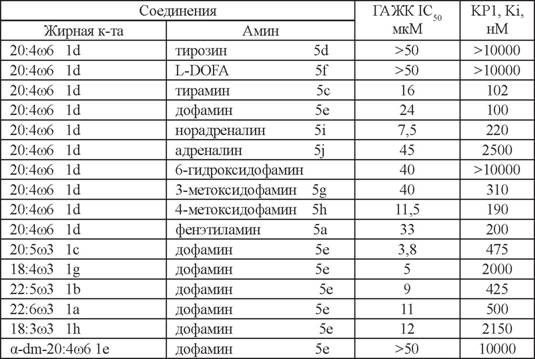
Таблица 3
Ингибирование ГАЖК N-ацил-серотонинами и их аналогами
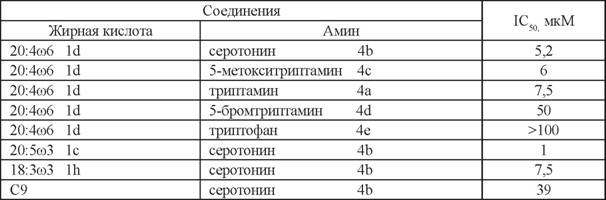
в ароматической части. Об этом свидетельствуют результаты по активности соединений, где эти группы отсутствовали или были замещены. Также наблюдалось значительное снижение активности у ацилированных производных аминокислот, которые являются предшественниками дофамина и серотонина: N-арахидоноилти- розина и N-арахидоноилтриптофана (см. табл. 2, 3).
Полученные данные дают основание заключить, что производные жирных кислот с дофамином и серотонином являются умеренными ингибиторами ГАЖК. Можно предположить, что повышение количества таких нейролипинов в результате их эндогенного выделения или поступления извне будет приводить к усилению физиологических ответов как за счет собственного взаимодействия с рецепторными мишенями, так и за счет торможения инактивации других нейролипинов, например анандамида.
4.
Еще по теме Влияние ацилированных производных дофамина и серотонина на гидролиз анандамида:
- Антиагрегационное действие амидов жирных кислот с дофамином и серотонином
- Д. Дофамин.
- IV. Производные имидазола
- Ферментативный гидролиз РНК
- Гидролиз РНК химическими агентами
- Е. 5-Гидрокситриптамин (серотонин).
- Производные оксина.
- Роль дофамина в регуляции цикла бодрствование-сон
- Определение содержания серотонина.
- 214. Поражения производными нитрофенолов и цианидами
- Нарушения гидролиза и всасывания углеводов
- Нарушения гидролиза и всасывания белков
- 2). Отравления транквилизаторами (производные бензодиазепина)
- Селективные ингибиторы обратного захвата серотонина и норадреналина (СИОЗСН)
- Селективные ингибиторы обратного захвата серотонина