Роль дофамина в регуляции цикла бодрствование-сон
C чем же связаны нарушения сна при БП? Ранние гипотезы связывали их, естественно, с разрушением продуцирующей ДА нигростриатной системы. Однако роль ДА в нормальной регуляции бодрствования и сна долгие десятилетия оставалась загадочной.
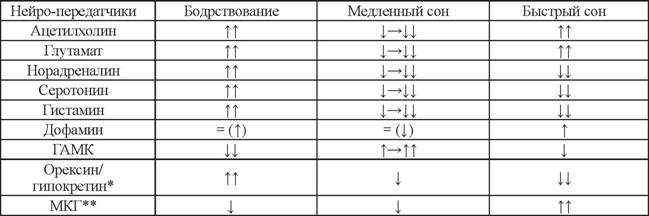
Другие аминергические системы мозга - норадренергическая (NA), клетки которой расположены в области синего пятна (LC), серотонинерги- ческая (5-HT), локализованная главным образом в дорзальных ядрах шва (DR), и гистаминергическая (HA), находящаяся в туберомаммиллярных ядрах заднего гипоталамуса (TMH), оказывают мощные активирующие (тонические деполяризующие) воздействия, как восходящие (на нейроны неокортекса и архипалеокор- текса), так и нисходящие (на мотонейроны спинного мозга). Нервные клетки этих систем, весьма активные в бодрствовании, прогрессивно снижают свою импульса- цию в медленном сне и полностью (или почти полностью) «замолкают» в быстром (табл. 1). Таким образом, эти три мозговые системы, наряду с холинергической (Ach), глутаматергической (Glu) и сравнительно недавно открытой орексин/гипок- ретинергической (Orx/Hcr), участвуют в поддержании «тонуса неокортекса» и мышечного тонуса в бодрствовании (Ковальзон, 2011а,б; Datta, 2010; Datta, MacLean, 2007; Pace-Schott, Hobson, 2002).Разрушение выделяющих дофамин нейронов vPAG у крыс приводит к увеличению продолжительности медленного и быстрого (парадоксального) сна за счет снижения суммарного времени бодрствования (Monti, Jantos, 2008), что позволило также отнести их к системе поддержания бодрствования. Более того, у мышей, но- каутных по гену дофаминового переносчика, с увеличенным содержанием внеклеточного дофамина, наблюдается повышенная представленность бодрствования и пониженная - медленного сна (примерно на 1/5) в светлый (неактивный) период суток по сравнению с гетерозиготными и контрольными (немутантными, wild-type) животными (Wisor et al., 2001).
Однако ранние исследования не выявили заметных изменений активности ДА нейронов, расположенных в SNpc/VTA, в цикле сон-бодрствование у крыс и кошек (Miller et al., 1983; Steinfels et al., 1983; Trulson et al., 1981; Trulson, Preussler, 1984). Поэтому долгое время считалось, что, в отличие от вышеперечисленных мозговых
Таблица 1
Упрощенная схема выделения основных медиаторов головного мозга в цикле бодрствование-сон
*Орексин (гипокретин) - открытый в 1998 г. пептид-медиатор ЦНС. **МКГ - меланин- концентрирующий гормон холоднокровных позвоночных, открытый в 1983 г; в 1985 г. была обнаружена роль пептида-медиатора в ЦНС млекопитающих.
Стрелка вверх - повышение выделения; двойная стрелка вверх - значительное повышение выделения; стрелка вниз - снижение выделения; двойная стрелка вниз - значительное снижение выделения; горизонтальная стрелка вправо - постепенное понижение/повышение выделения; знак равенства - выделение без изменений; стрелка в скобках - данные сомнительны.
аминергических систем, ДА-нейроны нигростриатной системы не участвует в регуляции бодрствования и сна. В дальнейшем, однако, было обнаружено, что концентрация внеклеточного ДА в местах проекции нигростриатных нейронов колеблется в цикле бодрствования-сна, снижаясь в медленном сне по сравнению с бодрствованием и вновь повышаясь в быстром (Lena et al., 2005). Оставалось непонятным, за счет чего же изменяется выброс ДА в стриатуме, если частота импульсации нейронов, его синтезирующих, не меняется? Однако более тщательное изучение характеристик ДА-нейронов VTA выявило существенные различия в рисунке (паттерне) разрядов в быстром сне по сравнению со спокойным бодрствованием и медленным сном (Dahan et al., 2007). В быстром сне значительно возрастает представленность разрядов в виде «вспышек», «пачек» (bursts), причем внутри каждой «вспышки» амплитуда разрядов прогрессивно снижается (рис. 2а,с). Аналогичные изменения рисунка разрядов ДА-нейронов VTA (нарастание представленности «пачек») отмечаются при переходе от спокойного бодрствования к эмоционально-мотивационному поведению с положительным подкреплением (например, поеданию крысой вкусной пищи, рис.
2b,d). Именно такие «вспышки» сопровождаются массивным выбросом ДА в синаптические щели и межклеточное пространство (Wightman, Robinson, 2002).Иммуногистохимическое исследование экспрессии c-Fos белка ДА клетками головного мозга крыс показало, что нейроны VTA, в отличие от SNpc, увеличивают свою активность при «отдаче» быстрого сна после его 2-суточной депривации (Maloney et al., 2002). Недавнее исследование французских авторов, однако, не выявило изменений Fos-экспрессии в областях А9 и А10 (SNpc/VTA и vPAG) во время восстановительного сна («отдачи») после 3-суточной депривации быстрого сна или 3-часового пребывания в бодрствовании в сенсорно обогащенной среде. Было обнаружено лишь небольшое повышение активности группы ДА нейронов, расположенных в каудальном отделе гипоталамуса (А11). Кроме того, было отмечено повышение количества Fos-иммунореактивных ДА-содержащих клеток, локализованных в zona incerta (A13) (Leger et al., 2010). Однако те же авторы отмечают, ссылаясь на работу (Kovacs, 1998), что с-Fos-метод не является надежным маркером активации нейронов (Luppi et al., 2011).
Надо сказать, что в обеих приведенных выше работах (Maloney et al., 2002; Leger et al., 2010) для депривации быстрого сна авторы использовали разработанный в свое время в лаборатории М. Жуве метод «малых площадок». Хорошо известно, однако, что этот поведенческий метод хотя и прост в использовании, вызывает не только эффективное подавление быстрого сна, но и значительное снижение - медленного, а также сильный эмоциональный стресс у подопытных животных (мышей, крыс, кошек), вызванный страхом упасть в воду во время сна. Недаром он был назван известным патофизиологом Ф.З. Меерсоном методом «стресса по Жуве»! При этом происходит взаимодействие стресса и депривации сна и формирование особого фенотипа, сохраняющегося на протяжении по крайней мере первых суток после завершения депривации. Все это делает данную методику неадекватной для изучения эффектов избирательного лишения быстрого сна «в чистом виде», а результаты, полученные с ее помощью, - не интерпретируемыми.
Для изучения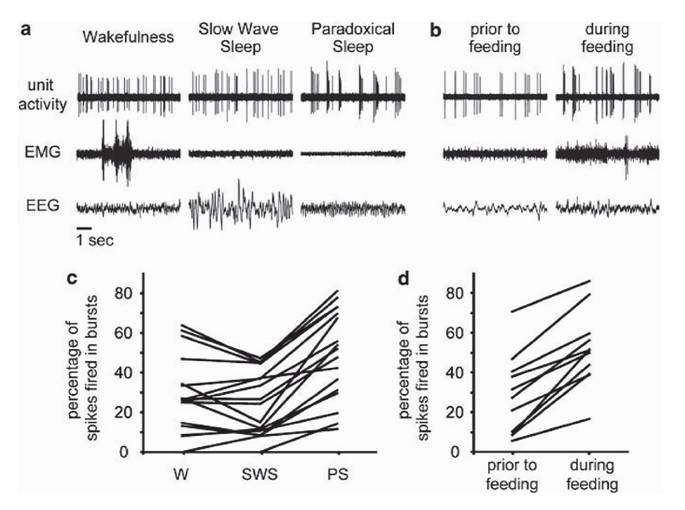
Рис. 2. ДА нейроны вентральной покрышки переходят в «пачечный» режим разрядов в быстром (парадоксальном) сне (а, c) и при потреблении крысой вкусной пиши (b, d). Вверху: разряды одиночных нейронов (unit activity), одного - в цикле бодрствование-сон (wakefulness, W - бодрствование, Б; slow wave sleep, SWS - медленноволновый сон, МС; paradoxical sleep, PS - парадоксальный сон, ПС), а другого - до- (prior to feeding) и во время (during feeding) поедания вкусной пиши. EMG - элек- тромиограмма (ЭМГ); EEG - электроэнцефалограмма (ЭЭГ).Б - характеризуется выраженной активностью на ЭМГ и низкоамплитудной, десинхронизированной ЭЭГ; МС - пониженной активностью на ЭМГ и высоковольтными медленными волнами на ЭЭГ; ПС - исчезновением мышечного тонуса на ЭМГ и выраженным тета-ритмом на ЭЭГ. Отметка времени - 1 с. ДА нейроны переключаются от нерегулярных разрядов с редкими дуплетами в спокойном бодрствовании и МС - к «пачечному» рисунку в ПС и при еде, причем каждая пачка состоит из нескольких спайков с прогрессивно снижающейся амплитудой. Также видно, что фазическая активация ЭМГ в Б не влияет на импульсацию. Внизу: представленность пачечной активности в процентах от всех зарегистрированных спайков (percentage of spikes fired in bursts) у 17 нейронов, зарегистрированных в цикле бодрствование-сон, и 11 нейронов, записанных до и во время поедания вкусной пищи. Видно, что представленность пачек нарастает во время ПС и еды у всей популяции ДА-нейронов. Между нейронной активностью в Б и МС достоверных различий не выявляется (Dahan et al, 2007), печатается с разрешения
эффектов депривации быстрого сна уже давно разработаны иные, гораздо более «деликатные» методы (Kovalzon, Tsibulsky, 1984; Ковальзон, Цибульский, 1985).
Какова же роль повышенного уровня дофамина в быстром сне? Обобщая свои наблюдения за больными с различными неврологическими нарушениями, британский нейропсихолог Марк Солмс писал о том, что у многих больных с поражениями ствола объективно регистрируемое подавление быстрого сна не сопровождается исчезновением субъективно переживаемых сновидений.
Наоборот, полное выпадение отчетов о сновидениях отмечается у тех больных, у которых поражения находятся в области, казалось бы, никакого отношения к регуляции быстрого сна не имеющей - вентромезиального лобного белого вещества (Solms, 2009). Однако именно в этой области проходит проекционный ДА-путь от VTA/SNpc к прилежащим ядрам перегородки (NAcc) и далее - к лобной коре (FC, рис. 3). Эта же область оказывается разрушенной при фронтальной лейкотомии, после которой у больных исчезают галлюцинации, бред, а заодно и сновидения.Наряду с нейрохирургическими были получены и психофармакологические доказательства важнейшей роли системы ДА в возникновении сновидений. Хорошо известно, что шизофреническая симптоматика связана с избыточной продукцией мозгового дофамина (наряду со снижением глутамата, норадреналина и серотонина) и лечится подавлением ДА передачи с помощью галоперидола и других анти- психотиков, подавляющих сновидения. Наоборот, недостаточность дофаминовой передачи, характерная для БП и вызывающая двигательные нарушения, приводит к полному прекращению отчетов больных о сновидениях (Lima et al., 2007). Лечится же БП ДА препаратами (леводопой и другими), прием которых, судя по отчетам больных, резко активирует переживание ими сновидений (Lena et al., 2004).
Таким образом, согласно точке зрения М. Солмса, быстрый сон и сновидения - явления связанные, в норме протекающие одновременно, но отнюдь не тождественные. Это коренным образом расходится с классической гипотезой М. Жуве, разработанной еще в 1960-е годы и представленной им не только в научной и научно-
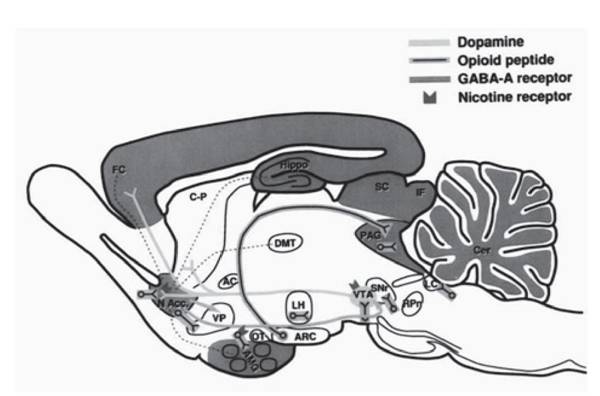
Рис. 3. Дофаминергическая система вентральной покрышки и ее проекции на парасаггиттальном срезе головного мозга крысы (SNr=SNpc), с разрешения (Roehrs, 2007)
популярной, но и в художественной литературе (Жуве, 2006, 2008). Если быстрый сон связан с активацией ромбэнцефалических и гипоталамических структур, использующих в качестве нейропередатчиков глутамат, ацетилхолин, ГАМК и МКГ, то материальная основа такого сложного психологического явления, каким является переживание эмоционально насыщенных сновидений, состоит, по М.
Солмсу, в активации ДА-ергических структур среднего и переднего мозга (Solms, 2009).Резюмируя, можно сказать, что ДА-ергические нейроны связаны с регуляцией цикла сон-бодрствование главным образом тем, что участвуют в поддержании эмоциональных проявлений бодрствования и быстрого сна. Соответственно, недостаточность ДА, постепенно развивающаяся в ходе нейродегенерации при БП, должна, видимо, проявляться в первую очередь как нарастание сонливости и ослабление выраженности сновидений. Кроме того, она может косвенно (через усиление ГАМК-ергического торможения холинергического «центра быстрого сна» в области PPT/LDT за счет активации ретикулярной части черной субстанции) вызывать и некоторое снижение представленности быстрого сна (урежение и/или укорочение его эпизодов), которое снимается ДА-препаратами (Takakusaki et al., 2004; Силькис, 2006). Однако приведенный выше перечень нарушений сна при БП гораздо шире.
4.
Еще по теме Роль дофамина в регуляции цикла бодрствование-сон:
- 65особ цикла сон бодрствов у детей
- 33. Сон и бодрствование. Физиологические механизмы влияния на здоровье.
- Блок регуляции тонуса и бодрствования
- Основы регуляции клеточного цикла
- Параграф седьмой. О заключениях по двигательным действиям [мозга] и по тому, что с ними сходно, как [например], сон и бодрствование
- 6.4. Сон и его регуляция.
- Роль холинергической системы стриатума в регуляции двигательного поведения
- Роль субталамического ядра в регуляции двигательного поведения
- 1.2. Роль вегетативной регуляции в формировании артериальной гипертензии
- Роль белков теплового шока в Т-клеточной регуляции хронического воспаления
- Роль слюны в питании и регуляции иммунных реакций хозяев
- Дофаминергическая система среднего мозга и ее роль в регуляции моторного поведения
- Блок программирования, регуляции и контроля сложных форм деятельности. Подкорковые образования мозга и их роль