Природа флуорофоров, ответственных за аутофлуоресценцию глазного дна
Как известно, фотолиз зрительного пигмента родопсина завершается разрывом ковалентной связи между опсином и полностью-транс-ретиналем (ПТР) и высвобождением последнего из хромофорного центра опсина.
ПТР высвобождается во внутридисковое пространство фоторецепторного диска наружного сегмента зрительной клетки. Затем он переносится через фоторецепторную мембрану на цитоплазматическую поверхность диска, где восстанавливается до полностью- транс-ретинола высокоактивной ретинолдегидрогеназой. В свою очередь, полно- стью-транс-ретинол переносится при помощи ретинол-переносящих белков из наружного сегмента палочки в клетку РПЭ (Palczewski et al., 1994).При патологии, когда белок-переносчик дефектен, например при болезни Штаргардта (дефектным является белок-переносчик ABCR4), ПТР накапливается в фоторецепторной мембране (Travis, 1998; Островский, 2005; Rozanowska, Sama, 2005).
При избыточном обесцвечивании родопсина в фоторецепторной мембране также может происходить накопление ПТР. Свободный и неудаленный из фоторецепторной мембраны ПТР может легко взаимодействовать с аминогруппой фосфатидилэтаноламина с образованием основания Шиффа - N-ретинилиден- фосфатидилэтаноламина (АРЕ). Именно этот продукт взаимодействия ПТР с фосфатидилэтаноламином удаляется из фоторецепторной мембраны при помощи АТФ-зависимого белка-переносчика (ABCR) (Weng et al., 1999; Saari, 2000; Lamb, Simon, 2004; Molday et al., 2009; Maeda et al., 2012).
Неудаленный АРЕ может вступать во взаимодействие еще с одной молекулой ПТР, приводя к образованию бисретиноидов, в том числе таких побочных и потенциально опасных продуктов, как N-бис-ретинилиден-фосфатидилэтаноламин - А2-РЕ (N-bis-retinylidene-phosphatidylethanolamine), дигидро-А2-РЕ, основания Шиффа между димером ПТР и фосфатидилэтаноламином - ATR-dimer-PE (ATR- dimer-phosphatidylethanolamine conjugate) (Liu et al., 2000; Kim et al., 2007).
При взаимодействии двух молекул ПТР с аминогруппами белка (родопсина) может образовываться А2-родопсин (Fishkin et al., 2003). Все эти побочные продукты являются устойчивыми соединениями, и они представляют собой источник накопления бисретиноидов в фоторецепторной клетке (Rozanowska, Sarna, 2005).В то же время, сам ПТР является эффективным природным фотосенсибилизатором (Harper, Gaillard, 2003; Fu et al., 2003; Liu, Hammand, 2003). По механизму фотосенсибилизированного окисления ПТР способен окислять и белки, и липиды в фоторецепторной мембране. Показано, что он способен повреждать родопсин, межфоторецепторный ретинол-связывающий белок (Островский, Федорович, 1996; Fedorovich et al., 2000), а также мембранный белок-переносчик ABCR (Sun, Nathans, 2001).
До недавнего времени считалось, что при физиологических условиях накопление токсичного и фототоксичного свободного ПТР в наружном сегменте зрительной клетки или не происходит, или является кратковременным. Однако совокупность накопленных к настоящему времени данных свидетельствует о том, что ПТР способен выступать в роли токсического или фототоксического агента, усугубляющего развитие дегенеративных заболеваний сетчатки (Rozanowska, Sama, 2005; Masutomi et al., 2012; Maeda et al., 2012). Другими словами, накопление в фоторецепторных клетках свободного ПТР может рассматриваться как фактор риска усугубляющего действия света при развитии различных дегенеративных заболеваний сетчатки (Островский, 2005; Wang et al., 2003).
В процессе эволюции зрительной системы сформировался достаточно эффективный способ защиты от накопления в клетке молекулярных дефектов и, в том числе, побочных продуктов фотолиза родопсина - бисретиноидов. Постоянное обновление фоторецепторных мембран и всего наружного сегмента зрительной клетки (и палочки, и колбочки) позволяет в норме избегать накопления молекулярных дефектов, благодаря чему молекулярная «машинерия» зрения остается эффективной на протяжении всей жизни. Однако вместе с т.н. фагосомами - «отработанными» и фагоцитированными обломками наружных сегментов фоторецепторных клеток, в клетки РПЭ попадают и (фото)токсичные бисретиноиды.
В результате неполной лизосомальной деградации фагосом в клетках РПЭ накапливаются неде- градабельные ЛГ - «пигмент старости».Долгое время ЛГ считались инертным побочным продуктом жизнедеятельности клетки РПЭ. Однако, во-первых, была показана способность ЛГ генерировать при действии видимого света активные формы кислорода (Островский и др., 1996; Boulton et al., 1993) и, во-вторых, выявлена явная корреляция между накоплением ЛГ в клетках РПЭ и дегенеративными заболеваниями сетчатки, в том числе ВМД (Boulton et al., 1993, 2004; Brunk, Terman, 2002; Островский, 2005). Не вызывает сомнения, что производные ПТР - бисретиноиды, являются фототоксичными для клеток РПЭ (Suter et al., 2000; Ben-Shabat et al., 2002а; Kim et al., 2006) и что их окисленные формы становятся токсичными и в темноте (Ben-Shabat et al., 2002а, Sparrow et al., 2003; Rozanowska, Sarna, 2005).
ЛГ поглощают свет в видимой, в основном коротковолновой (фиолетово-синей), области спектра, и обладают ярко выраженной флуоресценцией. Флуоресцентные свойства ЛГ обусловлены, в основном, наличием в них конъюгатов ПТР различной природы. Однако хорошо изученным является практически только один флуоро- фор, т.н. А2Е (К-ретинил-К-ретинилиденэтаноламин) - продукт взаимодействия двух молекул ПТР с аминогруппой одного из липидов фоторецепторной мембраны - фосфатидилэтаноламина (РЕ) (Parish et al., 1998; Ben-Shabat et al., 2002b). A2E образуется из своего предшественника А2РЕ путем ферментативного гидролиза (рис. 1). Помимо А2Е в ЛГ содержится еще около двадцати флуорофоров, являющихся производными ретиналя (Eldred, Katz, 1988; Sparrow et al., 2009). В настоящее время охарактеризованы такие конъюгаты ПТР, как ПТР-димер (ATR-dimer), ПТР-димер этаноламин конъюгат (ATR-dimer-E), ПТР-димер фосфа- тидилэтаноламин конъюгат (ATR-dimer-PE) (Kim et al., 2007), А2-дигидропиридин- фосфатидилэтаноламин (A2-DHP-PE) и А2-дигидропиридин-этаноламин (A2- DHP-E) (Wu et al., 2009), а также конъюгат ПТР с глицерофосфоэтаноламином (glycerophosphoethanolamine) (A2-GPE) (Yamamoto et al., 2011).
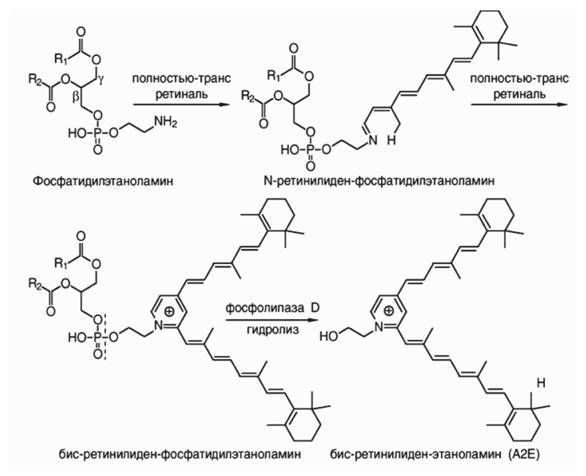
Рис. 1. Последовательность химических реакций, приводящих к образованию молекулы А2Е (і¥-ретинил-і¥-ретинилидеютаноламина) из двух молекул полностью-транс-ретиналя и молекулы PE (фос фатидилэтаноламина)
В присутствии кислорода А2Е может фотоокисляться с образованием различных окисленных форм (Avalle et al., 2004; Dillon et al., 2004). Окисленные продукты А2Е способны оказывать токсическое действие на клеточные структуры уже без участия света (Sparrow et al., 2003; Wang et al., 2006; Wu et al., 2010; Yoon et al., 2012). Эти продукты окисления являются более гидрофильными, чем исходный А2Е. Поэтому они могут высвобождаться из ЛГ (Соколов и др., 2005) и перемещаться в цитоплазме клетки РПЭ в другие клеточные органеллы, например в ядро и в митохондрии РПЭ (Донцов и др., 2009). Окисленные формы А2Е были обнаружены в ЛГ, выделенных как из старческих глаз человека, так и из РПЭ глаз мышей с мутацией в ABCR-гене, моделирующем болезнь Штаргарда и ВМД (Avalle et al., 2004; Radu et al., 2004, Wielgus et al., 2010). При этом были идентифицированы в РПЭ как мышей, так и человека обе формы продуктов окисления А2Е - эпокси- и фуранопроизводные (Jang et al., 2005; Яковлева и др., 2006). Важно подчеркнуть, что образование и накопление окисленных продуктов отражается на спектральных характеристиках ЛГ. Как нами ранее было показано, при действии света (фотоокислении) спектры поглощения и флуоресценции как А2Е, так и самих ЛГ сдвигаются в коротковолновую область (Донцов и др., 2005, 2006, 2009; Яковлева и др., 2006, 2009; Фельдман и др., 2010). Очевидно, что неокисленные и окисленные формы
флуорофоров могут вносить различный вклад в картину аутофлуоресценции глазного дна, что следует учитывать при дальнейшем усовершенствовании этого метода.
Изучение состава и спектральных характеристик флуорофоров, их неокисленных и окисленных форм, содержащихся как в ЛГ клеток РПЭ, так и в фоторецепторных клетках сетчатки, является важным как для понимания природы, так и для интерпретации картины аутофлуоресценции глазного дна, меняющейся в ходе старения и на различных стадиях патологического процесса. Такое изучение важно независимо от того, возбуждается АФ при 488 нм или регистрируется в диапазоне 500-600 нм, как это имеет место в существующем гейдельбергском офтальмоскопе, и это тем более важно, если в будущем удастся получать картину АФ при более коротковолновом возбуждении и более коротковолновой регистрации.
4.
Еще по теме Природа флуорофоров, ответственных за аутофлуоресценцию глазного дна:
- Флуорофоры липофусциновых гранул в клетках ретинального пигментного эпителия и аутофлуоресценция глазного дна человека как новый неинвазивный метод диагностики старческих изменений и дегенеративных заболеваний сетчатки
- Изменения глазного дна п р и с и ф и л и с е.
- Изменения глазного дна при токсикозах беременных
- Исследование глазного дна.
- 46. Изменения глазного дна при миопии
- Изменение глазного дна при туберкулезе.
- Изменения глазного дна при заболеваниях почек
- Спектральные характеристики флуорофоров липофусциновых гранул
- Выбор способа периоперационной антибиотикопрофилактики при глазных операциях со вскрытием глазного яблока
- Флуорофоры липофусциновых гранул из клеток ретинального пигментного эпителия кадаверных глаз человека
- Образование флуорофоров - продуктов взаимодействия свободного транс-ретиналя с аминогруппами белка и липидов в фоторецепторных клетках (исследование in vitro на модельных системах)
- Спектры флуоресценции и состав флуорофоров липофусциновых гранул ретинального пигментного эпителия кадаверных глаз в двух случаях визуализируемой абиотрофии
- Метод одновременного спектроскопического анализа эндогенных флуорофоров и 5-АЛК-индуцированного протопорфирина IX, содержащихся в нервных тканях
- ОФТАЛМОСКОПИЧЕСКАЯ КАРТИНА НОРМАЛЬНОГО ГЛАЗНОГО ДНА
- 18. Рак дна полости рта.
- ИССЛЕДОВАНИЕ СРЕД ГЛАЗА И ГЛАЗНОГО ДНА
- 16.Правосознание и ответственность личность.