оПХЛЕМЕМХЕ ЮДЕМНГХМТНЯТЮРНБ ДКЪ ОПЕДНРБПЮЫЕМХЪ ЛНКЕЙСКЪПМНЦН ЯРЮПЕМХЪ NADH-ДЕЦХДПНЦЕМЮГШ Б ЛХРНУНМДПХЪУ
л.я. тПНКНБЮ, м.к. бЕЙЬХМ
мЕИПНДЕЦЕМЕПЮРХБМШЕ ОПНЖЕЯЯШ Б ЛНГЦЕ ОПХБНДЪР, Б ВЮЯРМНЯРХ, Й ОЮПЙХМЯНМХГЛС. яРЮПВЕЯЙХИ ОЮПЙХМЯНМХГЛ Я РПСДНЛ ОНДДЮЕРЯЪ КЕВЕМХЧ. оПХЛЕМЕМХЕ ПЮГКХВМШУ ЛЕДХЙЮЛЕМРНБ ОНЛНЦЮЕР МЮ БПЕЛЪ ЯМЪРЭ ЯХЛОРНЛШ, МН МЕ АНКЕЕ РНЦН.
мЮОПХЛЕП, ДНТЮЛХМНБЮЪ РЕПЮОХЪ МЕ ОНГБНКЪЕР БШКЕВХРЭ ОЮПЙХМЯНМХГЛ. оНЩРНЛС МЕНАУНДХЛШ МНБШЕ ОНДУНДШ, ЙНРНПШЕ ЛНЦКХ АШ ОНГБНКХРЭ НЯСЫЕЯРБКЪРЭ ЩТТЕЙРХБМНЕ КЕВЕМХЕ.нДХМ ХГ ОСРЕИ КЕВЕМХЪ, МЮ МЮЬ БГЦКЪД, ЛНЦ АШ ГЮЙКЧВЮРЭЯЪ Б БНЯЯРЮМНБКЕМХХ ТСМЙЖХХ ЯРЮПЕЧЫХУ ЛХРНУНМДПХИ. бЕДЭ ЯРЮПЕМХЕ ЛХРНУНМДПХИ ЛНФЕР ОПХБНДХРЭ Й ДЕЦЕМЕПЮЖХХ МЕИПНМНБ. яРЮПШЕ ЛХРНУНМДПХХ - ОНЯРЛХРНУНМДПХХ - УЮПЮЙРЕПХГСЧРЯЪ СУСДЬЕМХЕЛ ПЮАНРШ ДШУЮРЕКЭМНИ ЖЕОХ, ЯМХФЕМХЕЛ ОПНДСЙЖХХ юрт, ОНЪБКЕМХЕЛ КХОНТСЯЖХМЮ (аЕКЪЙНБХВ, 1990).
б ОНЯКЕДМХЕ ЦНДШ ЯРЮПВЕЯЙНЕ ПЮГБХРХЕ МЕИПНЛШЬЕВМШУ ДЕЦЕМЕПЮРХБМШУ ГЮАНКЕБЮМХИ БЯЕ ВЮЫЕ ЯБЪГШБЮЧР Я МЮПСЬЕМХЪЛХ ПЮАНРШ NADH-ДЕЦХДПНЦЕМЮГМНЦН ЙНЛОКЕЙЯЮ I Б ЛХРНУНМДПХЪУ (Seo et al., 2004; Varghese et al., 2009) Х, ЙЮЙ ЯКЕДЯРБХЕ, ОНБШЬЕММШЛ НЙХЯКХРЕКЭМШЛ ЯРПЕЯЯНЛ ЙКЕРНЙ.
лШ ОПЕДОНКЮЦЮЕЛ, ВРН НДМХЛ ХГ ТЮЙРНПНБ, БШГШБЮЧЫХЛ ЩРХ МЮПСЬЕМХЪ, ЪБКЪЕРЯЪ ОНРЕПЪ ЯБЪГХ ЛЕФДС FMN Х NADH-ДЕЦХДПНЦЕМЮГНИ.
FMN МЮУНДХРЯЪ Б 50-ЙдЮ ЯСАЗЕДХМХЖЕ NADH-ДЕЦХДПНЦЕМЮГШ (ХЛЕММН ЩРЮ ЯСАЗЕДХМХЖЮ ХЛЕЕР ЮЙРХБМШИ ЖЕМРП ДКЪ ЯБЪГШБЮМХЪ NADH). FMN ЯБЪГЮМ Я ТЕПЛЕМРНЛ МЕЙНБЮКЕМРМН Х ОНЩРНЛС ОПХ ПЮГКХВМШУ БНГДЕИЯРБХЪУ КЕЦЙН БШОЮДЮЕР. щРН ЛНФЕР ОПНХЯУНДХРЭ ЯОНМРЮММН Я РЕВЕМХЕЛ БПЕЛЕМХ, ОПХ МЕАНКЭЬНЛ ОНБШЬЕМХХ РЕЛОЕПЮРСПШ, ОНД ДЕИЯРБХЕЛ ЯКЕДНБШУ ЙНКХВЕЯРБ (0,001%) ДЕРЕПЦЕМРНБ Х ДПСЦХУ БПЕДМШУ ТЮЙРНПНБ (яНЙНКНБЮ, бЕЙЬХМ, 2008; бЕЙЬХМ, 2009). б ПЕГСКЭРЮРЕ ЩКЕЙРПНМШ НР NADH МЕ ХДСР ДЮКЕЕ ОН ЩКЕЙРПНМ-РПЮМЯОНПРМНИ ЖЕОХ, Ю МЮОПЪЛСЧ БНЯЯРЮМЮБКХБЮЧР ЛНКЕЙСКЪПМШИ ЙХЯКНПНД ДН ЯСОЕПНЙЯХДЮ. оЕПЕОПНХГБНДЯРБН ЯСОЕПНЙЯХД-ЮМХНМЮ, ЯОНЯНАМНЦН ОПХБНДХРЭ Й ОЕПЕЙХЯМНЛС НЙХЯКЕМХЧ КХОХДНБ, Й ОНБПЕФДЕМХЧ АЕКЙНБ Х дмй, БШГШБЮЕР МЕХГАЕФМНЕ ЯРЮПЕМХЕ ЙЮЙ ЯЮЛХУ ЛХРНУНМДПХИ, РЮЙ Х ДПСЦХУ ЙКЕРНВМШУ НПЦЮМЕКК.
б УНДЕ ЩРНЦН ЛНФЕР ГЮОСЯЙЮРЭЯЪ ЛЕУЮМХГЛ ЦХАЕКХ ЙКЕРЙХ ОСРЕЛ ЮОНОРНГЮ ХКХ МЕЙПНГЮ, Б ГЮБХЯХЛНЯРХ НР ЙНКХВЕЯРБЮ Х ЯРЕОЕМХ ОНБПЕФДЕМХЪ ЛХРНУНМДПХИ.пЮМЕЕ Б МЮЬЕИ КЮАНПЮРНПХХ АШКН ОНЙЮГЮМН, ВРН ОПЕДНРБПЮРХРЭ ОНРЕПЧ ЯБЪГХ ЛЕФДС NADH-ДЕЦХДПНЦЕМЮГНИ Х ЛНКЕЙСКНИ FMN ЛНФМН Я ОНЛНЫЭЧ NAD, ЙНРНПШИ ЯРЮАХКХГХПСЕР ЯНАНИ ТЕПЛЕМР (яНЙНКНБЮ, бЕЙЬХМ, 2008; бЕЙЬХМ, 2009). рЮЙНЕ ДЕИЯРБХЕ NAD БШГБЮМН, БН-ОЕПБШУ, РЕЛ, ВРН НМ, ЪБКЪЪЯЭ ОПНДСЙРНЛ НЙХЯКЕМХЪ NADH, ХМЦХАХПСЕР ПЕЮЙЖХЧ ДЕЦХДПХПНБЮМХЪ Х, БН-БРНПШУ, МЮУНДЪЯЭ Б ЮЙРХБМНЛ ЖЕМРПЕ, ЛЕУЮМХВЕЯЙХ ЯРЮАХКХГХПСЕР ЮЙРХБМШИ ЖЕМРП, ВРН ОПЕОЪРЯРБСЕР БШБЮКХБЮМХЧ FMN.
яНЙПЮЫЕМХЪ: FAD - ТКЮБХМЮДЕМХММСЙКЕНРХДШ; FMN-H2 - БНЯЯРЮМНБКЕММШИ ТКЮБХМ.
й ЯНФЮКЕМХЧ, БНГЛНФМНЕ КЕВЕМХЕ ЛХРНУНМДПХЮКЭМШУ ЩМЖЕТЮКНЛХНОЮРХИ Я ХЯОНКЭГНБЮМХЕЛ NAD ГЮРПСДМХРЕКЭМН, РЮЙ ЙЮЙ НМ ОКНУН ОПНМХЙЮЕР Б ЙКЕРЙХ Х Б ЯЮЛХ ЛХРНУНМДПХХ. йПНЛЕ РНЦН, NAD ЛНФЕР РНПЛНГХРЭ ПЮАНРС NADH-ДЕЦХДПНЦЕМЮГШ, ЮКЙН- ЦНКЭДЕЦХДПНЦЕМЮГШ Х ДПСЦХУ NAD-ГЮБХЯХЛШУ ТЕПЛЕМРНБ.
яБНЕИ ГЮДЮВЕИ Б ЩРНИ ПЮАНРЕ ЛШ ОНЯРЮБХКХ ОПНРЕЯРХПНБЮРЭ МЕЙНРНПШЕ ОПХПНДМШЕ ЮМЮКНЦХ NAD МЮ ОПЕДЛЕР ЯРЮАХКХГЮЖХХ FMN Б NADH-ДЕЦХДПНЦЕМЮГЕ, ОПХВЕЛ, ВРН НЯНАЕММН БЮФМН, АЕГ ОНДЮБКЕМХЪ ТЕПЛЕМРЮРХБМНИ ЮЙРХБМНЯРХ. дКЪ ЩРНЦН АШК БШАПЮМ ПЪД ОПХПНДМШУ БЕЫЕЯРБ Я ОНУНФЕИ МЮ NAD УХЛХВЕЯЙНИ ТНПЛСКНИ: юрP, юDP, юлP, ЮДЕМХМ, ЮДЕМНГХМ, МХЙНРХМНБЮЪ ЙХЯКНРЮ Х МХЙНРХМЮЛХД.
вРНАШ СБЕКХВХРЭ ЯОНМРЮММШИ БШУНД FMN, ХГНКХПНБЮММШЕ ЛХРНУНМДПХХ ХМЙСАХПНБЮКХЯЭ ОПХ РЕЛОЕПЮРСПЕ 37╟я Б РЕВЕМХЕ ДБСУ ВЮЯНБ. дКЪ РНЦН ВРНАШ РЮЙХЕ БЕЫЕЯРБЮ, ЙЮЙ NAD, NADH, ЮДЕМХМ Х ЮДЕМНГХМ, КСВЬЕ ОПНМХЙЮКХ Б ЛХРНУНМДПХХ, АШКЮ БШАПЮМЮ ЦХОНРНМХВЕЯЙЮЪ ЯПЕДЮ ХМЙСАЮЖХХ, ЯНДЕПФЮЫЮЪ 100 Лл ЯЮУЮПНГШ Х 10 Лл Tris-ТНЯТЮР, Пм=7,0. б РЮЙНИ ЯПЕДЕ ЛХРНУНМДПХЮКЭМШЕ ЛЕЛАПЮМШ ЯРЮМНБЪРЯЪ АНКЕЕ ОПНМХЖЮЕЛШЛХ. йПНЛЕ РНЦН, ЛХРНУНМДПХХ ХЯОНКЭГНБЮКХЯЭ ОНЯКЕ НДМНЙПЮРМНИ ГЮЛНПНГЙХ, ВРН ЯОНЯНАЯРБСЕР МЕЙНРНПНИ ОЕПЛЕЮАХКХГЮЖХХ ЛЕЛАПЮМ Х КСВЬЕЛС ОПНМХЙМНБЕМХЧ БЕЫЕЯРБ (МН МЕ ПЮГПСЬЮЕР ЯЮЛХУ НПЦЮМЕКК).
дКЪ НОПЕДЕКЕМХЪ ТЕПЛЕМРЮРХБМНИ ЮЙРХБМНЯРХ NADH-ДЕЦХДПНЦЕМЮГШ АШК БШАПЮМ ТНРНЛЕРПХВЕЯЙХИ ЛЕРНД аЕКЪЙНБХВЮ Я ОНЛНЫЭЧ ОЮПЮ-МХРПНРЕРПЮГНКХЪ ТХНКЕРНБНЦН (p-NTV), БНЯЯРЮМЮБКХБЮЧЫЕЦНЯЪ ДН ТНПЛЮГЮМЮ, ЙНРНПШИ НРЙКЮДШБЮЕРЯЪ Б ЛЮРПХЙЯЕ ЛХРНУНМДПХИ Б БХДЕ ЛХЙПНЙПХЯРЮККХЙНБ Х МЕ БКХЪЕР МЮ ДЮКЭМЕИЬСЧ ЯЙНПНЯРЭ ПЕЮЙЖХХ (аЕКЪЙНБХВ, 1990).
йПНЛЕ РНЦН, ЩРНР ЛЕРНД СДНАЕМ РЕЛ, ВРН ЯЙНПНЯРЭ БНЯЯРЮМНБКЕМХЪ p-NTV Б ДЕЯЪРЙХ ПЮГ ОПЕБНЯУНДХР ЯЙНПНЯРЭ ОЕПЕДЮВХ ЩКЕЙРПНМНБ ОН ЩКЕЙРПНМ-РПЮМЯОНПРМНИ ЖЕОХ МЮ ЙХЯКНПНД, ОНЩРНЛС ЛНФМН МЕ ХЯОНКЭГНБЮРЭ АКНЙЮРНПШ ДШУЮРЕКЭМНИ ЖЕОХ.лХРНУНМДПХЮКЭМСЧ ТПЮЙЖХЧ ХГ ОЕВЕМХ ЙПШЯ БШДЕКЪКХ Б ЯПЕДЕ, ЯНДЕПФЮЫЕИ 20 Лл Tris-HCl Х 250 Лл ЯЮУЮПНГШ (Пм=7,7), ОПХ РЕЛОЕПЮРСПЕ 2╟я ЯРЮМДЮПРМШЛ ЛЕРНДНЛ Я МЕЙНРНПШЛХ ЛНДХТХЙЮЖХЪЛХ (ьЮПНБЮ, бЕЙЬХМ, 2004). яМЮВЮКЮ ЖЕМРПХТСЦХПНБЮКХ ХЯУНДМСЧ НАЫСЧ ЯСЯОЕМГХЧ МЮ BECKMANJ2-21 ОПХ 1000 g Б РЕВЕМХЕ 10-15 ЛХМ, Б ПЕГСКЭРЮРЕ ВЕЦН НЯЮФДЮКХЯЭ (Х ГЮРЕЛ НРАПЮЯШБЮКХЯЭ) ЙКЕРЙХ Х ДПСЦХЕ ЙПСОМШЕ ВЮЯРХЖШ. яСОЕПМЮРЮМР ЖЕМРПХТСЦХПНБЮКХ Б РЕВЕМХЕ 15 ЛХМ ОПХ 5000 g, Б ПЕГСКЭРЮРЕ ВЕЦН НЯЮФДЮКХЯЭ Б НЯМНБМНЛ ГПЕКШЕ РЪФЕКШЕ ЛХРНУНМДПХХ, ЙНРНПШЕ ГЮРЕЛ ЯСЯОЕМДХПНБЮКХ Б РНИ ФЕ ЯПЕДЕ (9 ЛК МЮ 1.5 ЛК НЯЮДЙЮ), ПЮЯТЮЯНБШБЮКХ МЮ КЭДС Б ОПНАХПЙХ Eppendorf ОН 1 ЛК, ГЮЛНПЮФХБЮКХ Х ХЯОНКЭГНБЮКХ ДКЪ НОШРНБ (ОНЯКЕ ПЮГЛНПЮФХБЮМХЪ). б ПЪДЕ ЯКСВЮЕБ БШДЕКЕМХЕ ЛХРНУНМДПХИ ОПНХГБНДХКХ Б ЯПЕДЕ, ЯНДЕПФЮЫЕИ 200 Лл ЯЮУЮПНГШ, 100 Лл ЛЮММХР, 10 Лл УЕОЕЯ (Пм=7,5), Х НЯСЫЕЯРБКЪКХ ДБНИМСЧ ОПНЛШБЙС Я ДБСЙПЮРМШЛ ЖЕМРПХТСЦХПНБЮМХЕЛ ОПХ 5000 g.
тЕПЛЕМРЮРХБМЮЪ ПЕЮЙЖХЪ НЙХЯКЕМХЪ NADH ОЮПЮМХРПНРЕРПЮГНКХЕЛ ТХНКЕРНБШЛ (p-NTV) ДЕРЕЙРХПНБЮКЮЯЭ ОН НАПЮГНБЮМХЧ ТНПЛЮГЮМЮ, НРЙКЮДШБЮЧЫЕЦНЯЪ БМСРПХ ЛХРНУНМДПХИ. б УНДЕ ТЕПЛЕМРЮРХБМНИ ПЕЮЙЖХХ НЙХЯКЕМХЪ NADH ЛНКЕЙСКЮ p-NTV БНЯЯРЮМЮБКХБЮЕРЯЪ ДН ТНПЛЮГЮМЮ, Х ЯСЯОЕМГХЪ ЛХРНУНМДПХИ ОПХНАПЕРЮЕР ХГ- ГЮ МЕЦН ПНГНБСЧ НЙПЮЯЙС. б ЩРХУ НОШРЮУ Й ЯСЯОЕМГХХ ЛХРНУНМДПХИ ХГ ОЕВЕМХ ЙПШЯШ ДНАЮБКЪКХ 100 ЛЙл NADH, 100 ЛЙл p-NTV Х РЕЯРХПСЕЛНЕ БЕЫЕЯРБН (ADP, ЮДЕМНГХМ Х ДП.) Б ХГАШРЙЕ (ДН 300 ЛЙл). йХМЕРХЙС СБЕКХВЕМХЪ НОРХВЕЯЙНИ ОКНРМНЯ- РХ ОПХ НАПЮГНБЮМХХ ТНПЛЮГЮМЮ ХГЛЕПЪКХ Б РЕВЕМХЕ 15 ЛХМСР МЮ ЯОЕЙРПНТНРНЛЕРПЕ ╚оПНЛщЙНкЮА-5400ст╩ (яЮМЙР-оЕРЕПАСПЦ) ОПХ 540 МЛ.
йНМЖЕМРПЮЖХЧ ЛХРНУНМДПХЮКЭМНЦН АЕКЙЮ НОПЕДЕКЪКХ ст-ЩЙЯОПЕЯЯ-ЛЕРНДНЛ (бЕЙЬХМ, 1988). бЯЕ ТПЮЙЖХХ БШПЮБМХБЮКХ ОН ЯНДЕПФЮМХЧ АЕКЙЮ Я ОНЛНЫЭЧ АСТЕПЮ (100 Лл ЯЮУЮПНГШ Х 10 Лл Tris-ТНЯТЮР, Пм=7,0) ДН ЙНМЕВМНИ ЙНМЖЕМРПЮЖХХ 0,3 ЛЦ/ЛК.
яОЕЙРПШ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ ЛХРНУНМДПХИ ГЮОХЯШБЮКХ МЮ ЯОЕЙРПНТКС- НПХЛЕРПЕ SLM-4800 (SLM, Inc., яью) Б 1-ЯЮМРХЛЕРПНБНИ ХКХ 0,4-ЯЮМРХЛЕРПНБНИ ЙЧБЕРЮУ ОПХ 20╟я.
дКХМЮ БНКМШ БНГАСФДЕМХЪ ТКЮБХМНБ АШКЮ Б ЛЮЙЯХЛСЛЕ ОНКНЯШ ОНЦКНЫЕМХЪ ОПХ 450 МЛ, Ю ДКХМЮ БНКМШ ХГКСВЕМХЪ - Б ЛЮЙЯХЛСЛЕ ОПХ 525 МЛ. б ПЪДЕ НОШРНБ, ЦДЕ ХМРЕМЯХБМНЯРЭ ТКСНПЕЯЖЕМЖХХ АШКЮ МХГЙНИ, Б ВЮЯРМНЯРХ - ОПХ ХГЛЕПЕМХХ ЯРЕОЕМХ ОНКЪПХГЮЖХХ (P) ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ, ХЯОНКЭГНБЮКХ ГЕПЙЮКЭМШЕ ЙЧБЕРШ, ОНГБНКЪЧЫХЕ СЯХКХРЭ ЯХЦМЮК Б МЕЯЙНКЭЙН ПЮГ (бЕЙЬХМ, 2009).дКЪ НАМЮПСФЕМХЪ КНЙЮКХГЮЖХХ ТКЮБХМЮ ОНЯКЕ 2-ВЮЯНБНИ ХМЙСАЮЖХХ ОПХ 37╟я ПЮЯРБНП ЛХРНУНМДПХИ ОПНОСЯЙЮКЯЪ ВЕПЕГ 0,2-ЛХЙПНММШЕ ЛХККХОНПНБШЕ ТХКЭРПШ Х ГЮРЕЛ ОПНБНДХКНЯЭ ХГЛЕПЕМХЕ ТКСНПЕЯЖЕМЖХХ ТХКЭРПЮРЮ (ЛХРНУНМДПХХ НЯЕДЮКХ МЮ ТХКЭРПЕ). вЕПЕГ РЮЙХЕ ОНПШ ОПНУНДЪР ОНЛХЛН ТКЮБХМНБ Х ТКЮБНАЕКЙНБ РЮЙФЕ ОПНРНЛХРНУНМДПХХ (ЛЮКШЕ ГЮПНДШЬЕБШЕ МЕГПЕКШЕ ЛХРНУНМДПХХ), МН НМХ ОНВРХ МЕ ЯНДЕПФЮР ТКЮБХМНБ (гХМХМЮ, бЕЙЬХМ, 2008; бЕЙЬХМ, 2010). б ДПСЦХУ ХГЛЕПЕМХЪУ, ЦДЕ ЛХРНУНМДПХХ МЕ ЛЕЬЮКХ ХГЛЕПЕМХЧ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ, НМХ МЕ СДЮКЪКХЯЭ ХГ ПЮЯРБНПЮ. бЯЕ ХГЛЕПЕМХЪ ОПНБНДХКХЯЭ ОН 5-10 ПЮГ МЮ ЛХРНУНМДПХЮКЭМШУ ТПЮЙЖХЪУ ХГ ПЮГМШУ ФХБНРМШУ, ГЮРЕЛ МЮУНДХКНЯЭ ЯПЕДМЕЮПХТЛЕРХВЕЯЙНЕ ГМЮВЕМХЕ, ЙНРНПНЕ ГЮРЕЛ МНПЛХПНБЮКНЯЭ МЮ 100%.
пЮМЕЕ Б МЮЬЕИ КЮАНПЮРНПХХ АШКН ОНЙЮГЮМН, ВРН ОПНЖЕЯЯ БШУНДЮ FMN ХГ ЮЙРХБМНЦН ЖЕМРПЮ NADH-ДЕЦХДПНЦЕМЮГШ ЛНФЕР СЯЙНПЪРЭЯЪ МЕЙНРНПШЛХ БЕЫЕЯРБЮЛХ, Б ВЮЯРМНЯРХ - ЯКЕДНБШЛХ ЙНКХВЕЯРБЮЛХ ДЕРЕПЦЕМРНБ, Ю РЮЙФЕ МЕАНКЭЬХЛ ДКХРЕКЭМШЛ МЮЦПЕБЮМХЕЛ. щРН ЯНЦКЮЯСЕРЯЪ Я ДЮБМН ХГБЕЯРМШЛ ТЮЙРНЛ ОНКМНИ ХКХ ВЮЯРХВМНИ СРПЮРШ FMN Б УНДЕ ОПЕОЮПЮРХБМШУ ПЮАНР ОН БШДЕКЕМХЧ Х НВХЯРЙЕ ХГНКХПНБЮММНИ NADH-ДЕЦХДПНЦЕМЮГШ (Galante, Hatefi, 1979; Dooijewaard et al., 1978; Almeida et al., 1999; Cremona, Kerney, 1964; Okun et al., 2000). оПХВЕЛ СРПЮРЮ FMN ЯНОПНБНФДЮЕРЯЪ ОЮДЕМХЕЛ NADH:САХУХМНМ-ПЕДСЙРЮГМНИ ЮЙРХБМНЯРХ, МН МЕ NADH:ТЕППХЖХЮМХД- ПЕДСЙРЮГМНИ. щРН ЯБЪГЮМН Я РЕЛ, ВРН FMN БН БРНПНИ ПЕЮЙЖХХ МЕ СВЮЯРБСЕР (бЕЙЬХМ, 2009; ьЮПНБЮ, бЕЙЬХМ, 2004). мЕ СВЮЯРБСЕР НМ Х Б NADH:РЕРПЮГНКХИ-ПЕДСЙРЮГМНИ ПЕЮЙЖХХ (бЕЙЬХМ, 2009).
б МЮЬХУ ЩЙЯОЕПХЛЕМРЮУ ОН ЯРЮАХКХГЮЖХХ ТКЮБХМНБШУ ЛНКЕЙСК Б ТЕПЛЕМРЕ ЛШ ХМЙСАХПНБЮКХ ЛХРНУНМДПХХ Б РЕВЕМХЕ 2 ВЮЯНБ ОПХ 37╟я. оНЯКЕ ЩРНЦН ОПХ 20╟я ХГЛЕПЪКХ ТКЮБХМНБСЧ ТКСНПЕЯЖЕМЖХЧ (ЙБНГА=450 МЛ, ^ЩЛХЯЯ=525 МЛ).
йПНЛЕ РНЦН, ЛШ ХГЛЕПЪКХ РПХОРНТЮМНБСЧ ТКСНПЕЯЖЕМЖХЧ (ЙБНГА=286 МЛ, ^ЩЛХЯЯ=340 МЛ) - ДКЪ НОПЕДЕКЕМХЪ БШУНДЮ ХГ ЛХРНУНМДПХИ АЕКЙНБ (АНКЭЬХМЯРБН АЕКЙНБ ЛХРНУНМДПХИ ЯНДЕПФЮР РПХОРНТЮМНБШЕ НЯРЮРЙХ) Х ТКСНПЕЯЖЕМЖХЧ 7-ЮЛХМН-ЮЙРХМНЛХЖХМЮ (ЙБНГА=530 МЛ, ЙЩЛХЯЯ=620 МЛ) - ДКЪ НОПЕДЕКЕМХЪ БШУНДЮ дмй Б ПЮЯРБНП (7-ЮЛХМН-ЮЙРХМНЛХЖХМ УНПНЬН БЯРПЮХБЮЕРЯЪ Б ПЮЯОКЕРЕММШЕ СВЮЯРЙХ дмй) (бЕЙЬХМ, 2009). нЙЮГЮКНЯЭ, ВРН Б ОПНЖЕЯЯЕ БШУНДЮ FMN МЕ ОПНХЯУНДХР МХЙЮЙНЦН БШУНДЮ АЕКЙНБ Х дмй ХГ ЛХРНУНМДПХИ Б ПЮЯРБНП: ХМРЕМЯХБМНЯРЭ РПХОРНТЮМНБНИ ТКСНПЕЯЖЕМЖХХ Х 7-ЮЛХМН-ЮЙ- РХМНЛХЖХМНБНИ ТКСНПЕЯЖЕМЖХХ Б ЯПЕДЕ ОНЯКЕ 2-ВЮЯНБНИ ХМЙСАЮЖХХ ЛХРНУНМДПХИ (ОПХ 37╟я) Х ХУ ОНЯКЕДСЧЫЕЦН СДЮКЕМХЪ НЯРЮБЮКЮЯЭ ОПЕМЕАПЕФХЛН ЛЮКНИ (ДЮММШЕ МЕ ОПХБНДЪРЯЪ). щРН НГМЮВЮЕР, ВРН ЛХРНУНМДПХЮКЭМШЕ ЛЕЛАПЮМШ НЯРЮКХЯЭ ЖЕКШЛХ.оПХ ЩРНЛ ХМРЕМЯХБМНЯРЭ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ Б ЙНМРПНКЭМШУ ОПНАЮУ ОПХ ХМЙСАЮЖХХ БНГПЮЯРЮКЮ Б 2-3 ПЮГЮ (Б ПЮГМШУ НОШРЮУ, Б ГЮБХЯХЛНЯРХ НР ЯНУПЮММНЯРХ ЛХРНУНМДПХИ). мСФМН НРЛЕРХРЭ, ВРН БЙКЮД Б НАЫСЧ ТКЮБХМНБСЧ ТКСНПЕЯЖЕМЖХЧ ЛХРНУНМДПХЮКЭМНИ ЯСЯОЕМГХХ ДЮЕР МЕ РНКЭЙН FMN, ОПХМЮДКЕФЮЫХИ NADH-ДЕЦХДПНЦЕМЮГЕ, МН Х ТКЮБХМЮДЕМХММСЙКЕНРХДШ (FAD), ОПНВМН ЙНБЮКЕМРМН ЯБЪГЮММШЕ Я ДПСЦХЛХ ЛХРНУНМДПХЮКЭМШЛХ ДЕЦХДПНЦЕМЮГЮЛХ. хГЛЕМЕМХЕ ФЕ ХМРЕМЯХБМНЯРХ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ НАСЯКНБКЕМН Б НЯМНБМНЛ ХЛЕММН БШУНДНЛ FMN ХГ NADH-ДЕЦХДПНЦЕМЮГШ Б ПЮЯРБНП (ГЮ ЯВЕР БШУНДЮ FMN ХГ ЛХРНУНМДПХИ БНГПЮЯРЮЕР ЕЦН ЙНЩТТХЖХЕМР ЩЙЯРХМЙЖХХ, Ю ГМЮВХР Х ТКСНПЕЯЖЕМЖХЪ). лНКЕЙСКШ FAD ЛХРНУНМДПХЮКЭМШУ ТКЮБНОПНРЕХМНБ МЕ БМНЯЪР ГЮЛЕРМНЦН БЙКЮДЮ Б ХГЛЕМЕМХЕ ТКСНПЕЯЖЕМЖХХ, ОНРНЛС ВРН ЛНКЕЙСКШ FAD НЯРЮЧРЯЪ ЯБЪГЮММШЛХ ЙНБЮКЕМРМН ЯН ЯБНХЛХ ТЕПЛЕМРЮЛХ БМСРПХ ЛХРНУНМДПХЮКЭМШУ ЯТЕП, ПЮЯЯЕХБЮЧЫХУ ОЮДЮЧЫХИ МЮ МХУ ЯБЕР. й РНЛС ФЕ, ТКСНПЕЯЖЕМЖХЪ FAD ГЮРСЬЕМЮ ЮПНЛЮРХВЕЯЙХЛХ ЮЛХМНЙХЯКНРЮЛХ Х ФЕКЕГН-ЯЕПМШЛХ ЙКЮЯРЕПЮЛХ (бЕЙЬХМ, 2009).
оНЛХЛН ПЕЦХЯРПЮЖХХ ХГЛЕМЕМХИ ХМРЕМЯХБМНЯРХ ТКСНПЕЯЖЕМЖХХ ЛШ ХГЛЕПЪКХ ЯРЕОЕМЭ ОНКЪПХГЮЖХХ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ, ОН БЕКХВХМЕ ЙНРНПНИ ЛНФМН НОПЕДЕКХРЭ, Б ЙЮЙНЛ ЯНЯРНЪМХХ МЮУНДХРЯЪ FMN - ЯБЪГЮММНЛ ХКХ ЯБНАНДМНЛ (бЕЙЬХМ, 2009).
с ЯБЪГЮММНЦН ТКЮБХМЮ ЯРЕОЕМЭ ОНКЪПХГЮЖХХ БШЯНЙЮЪ (ХГ-ГЮ ЛЮКНЦН БПЕЛЕМХ ФХГМХ БНГАСФДЕММНЦН ЯНЯРНЪМХЪ Х МХГЙНИ БПЮЫЮРЕКЭМНИ ОНДБХФМНЯРХ БМСРПХ АЕКЙЮ). оН ЛЕПЕ БШУНДЮ FMN Б ПЮЯРБНП ЯРЕОЕМЭ ОНКЪПХГЮЖХХ СЛЕМЭЬЮЕРЯЪ, Р.Й. Б БНДМНИ ТЮГЕ ПЕГЙН СБЕКХВХБЮЕРЯЪ ЯЙНПНЯРЭ БПЮЫЕМХЪ, Ю РЮЙФЕ СБЕКХВХБЮЕРЯЪ БПЕЛЪ ФХГМХ (ХГ-ГЮ НРЯСРЯРБХЪ РСЬЕМХЪ ФЕКЕГН-ЯЕПМШЛХ ЙКЮЯРЕПЮЛХ). б МЮЬХУ НОШРЮУ ЯРЕОЕМЭ ОНКЪПХГЮЖХХ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ ОНЯКЕ ХМЙСАЮЖХХ ОПХ 37╟я ВЕПЕГ 2 ВЮЯЮ СЛЕМЭЬЮКЮЯЭ Б 1,5-2 ПЮГЮ.дКЪ РНЦН ВРНАШ ОПНБЕПХРЭ, ЦДЕ ХЛЕММН НЙЮГЮКЯЪ БШЯБНАНФДЕММШИ FMN - Б ЛХРНУНМДПХЮКЭМНЛ ЛЮРПХЙЯЕ ХКХ БМЕЬМЕЛ ПЮЯРБНПЕ - ЛШ ХГЛЕПЪКХ ХМРЕМЯХБМНЯРЭ Х ЯРЕОЕМЭ ОНКЪПХГЮЖХХ РЮЙФЕ Б ТХКЭРПЮРЮУ, ОНКСВЕММШУ ОСРЕЛ ТХКЭРПЮЖХХ ЯСЯОЕМГХИ ЛХРНУНМДПХИ (ДН Х ОНЯКЕ 2-ВЮЯНБНИ ХМЙСАЮЖХХ) ВЕПЕГ 0,2-ЛХЙПНММШЕ ЛХККХОН- ПНБШЕ ТХКЭРПШ. хМРЕМЯХБМНЯРЭ ТКСНПЕЯЖЕМЖХХ Б ТХКЭРПЮРЕ СБЕКХВХБЮКЮЯЭ ОНЯКЕ ХМЙСАЮЖХХ ЛХРНУНМДПХИ (Б ЯПЮБМЕМХХ Я ЛХРНУНМДПХЪЛХ ДН ХМЙСАЮЖХХ) Б 4,3 ПЮГЮ. щРН ЯНЯРЮБХКН 94% НР НАЫЕИ ХМРЕМЯХБМНЯРХ ТКСНПЕЯЖЕМЖХХ ОНЯКЕ ХМЙСАЮЖХХ. оПХ ЩРНЛ ЯРЕОЕМЭ ОНКЪПХГЮЖХХ ТКСНПЕЯЖЕМЖХХ Б ТХКЭРПЮРЕ ЯМХФЮКЮЯЭ НВЕМЭ ЯХКЭМН - ДН 0,06, ВРН ЯННРБЕРЯРБСЕР ЯБНАНДМНЛС БПЮЫЕМХЧ FMN Б БНДМНИ ТЮГЕ (бЕЙЬХМ, 2009). оНКСВЕММШЕ ДЮММШЕ НДМНГМЮВМН ЦНБНПЪР Н РНЛ, ВРН ОНВРХ БЕЯЭ FMN ОНОЮДЮЕР БН БМЕЬМХИ ПЮЯРБНП, ОПХВЕЛ Б ЯБНАНДМНЛ БХДЕ. лНКЕЙСКШ FMN, НРНЬЕДЬХЕ Б NADH-ДЕЦХДПНЦЕМЮГЕ НР ФЕКЕГН-ЯЕПМШУ ЙКЮЯРЕПНБ (ЙНРНПШЕ РСЬХКХ ТКСНПЕЯЖЕМЖХЧ FMN) Х БШЬЕДЬХЕ ХГ ТЕПЛЕМРЮ Б ЛЮРПХЙЯ (МН МЕ СЯОЕБЬХЕ БШИРХ Б МЮПСФМШИ ПЮЯРБНП), ДЮЧР Б ХГЛЕМЕМХЕ ХМРЕМЯХБМНЯРХ МЕАНКЭЬНИ БЙКЮД.
мСФМН НЯНАН ОНДВЕПЙМСРЭ, ВРН Б МЮЬХУ НОШРЮУ ХГЛЕМЕМХЪ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ МЕ ЯБЪГЮМШ Я ОПНЖЕЯЯЮЛХ НЙХЯКЕМХЪ/БНЯЯРЮМНБКЕМХЪ ТКЮБХМНБ, Р.Й. ДБСУВЮЯНБЮЪ ХМЙСАЮЖХЪ ЛХРНУНМДПХИ Б АНКЭЬХМЯРБЕ ЩЙЯОЕПХЛЕМРНБ НЯСЫЕЯРБКЪКЮЯЭ Б НРЯСРЯРБХЕ NADH, ЯСЙЖХМЮРЮ Х ХМШУ БНЯЯРЮМНБХРЕКЕИ ХКХ НЙХЯКХРЕКЕИ. рНКЭЙН Б НДМНЛ ЯКСВЮЕ - ОПХ ХЯОНКЭГНБЮМХХ NADH Б УНДЕ ХМЙСАЮЖХХ - ХЛЕКХ ЛЕЯРН НДМНБПЕЛЕММН ДБЮ ЪБКЕМХЪ: БНЯЯРЮМНБКЕМХЕ FMN ДН МЕТКСНПЕЯЖХПСЧЫЕЦН FMN-H2 Х ЯРЮАХКХГЮЖХЪ FMN Б ТЕПЛЕМРЕ. оНЯЙНКЭЙС БНЯЯРЮМНБКЕММШИ ТКЮБХМ (FMN-H2) ХЛЕЕР ПЮГПШУКЪЧЫХЕ НПАХРЮКХ Х МЕ ЯРЮАХКЕМ (ЯПЮГС НРДЮЕР ЩКЕЙРПНМШ Б ДШУЮРЕКЭМСЧ ЖЕОЭ, МЮ САХУХМНМ), РН НМ АШЯРПН ОЕПЕУНДХР БМНБЭ Б ЯБНЧ НЙХЯКЕММСЧ ТКСНПЕЯЖХПСЧЫСЧ ТНПЛС. рЮЙХЛ НАПЮГНЛ, БПЪД КХ ПЕДНЙЯ-ОЕПЕУНДШ FMN/FMN-H2 БМНЯЪР НЫСРХЛШИ БЙКЮД Б ХГЛЕМЕМХЪ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ.
лШ ОПНРЕЯРХПНБЮКХ ПЪД БЕЫЕЯРБ Я ВЮЯРХВМН ОНУНФЕИ МЮ NAD УХЛХВЕЯЙНИ ТНПЛСКНИ: ЮДЕМХМ, ЮДЕМНГХМ, ATP, ADP, AMP, МХЙНРХМНБЮЪ ЙХЯКНРЮ, МХЙНРХМЮЛХД, NAD.
мЮВМЕЛ Я ЮМЮКНЦНБ ЮДЕМХКНБНИ ВЮЯРХ ЙНТЕЛЕМРЮ: ЮДЕМХМ, ЮДЕМНГХМ, ATP, ADP, AMP.
оПХ ДНАЮБКЕМХХ ATP, ADP ХКХ AMP Б ЙНМЖЕМРПЮЖХХ 300 Лл Й ЯСЯОЕМГХХ ОЕВЕМНВМШУ ЛХРНУНМДПХИ ТКЮБХМНБЮЪ ТКСНПЕЯЖЕМЖХЪ ХГЛЕМЪКЮЯЭ, ЯННРБЕРЯРБЕММН, МЮ 44,8%, 49,4% Х 41% ЛЕМЭЬЕ ОН ЯПЮБМЕМХЧ Я ЙНМРПНКЕЛ (ПХЯ. 1, РЮАК. 1). оПХ ДН-
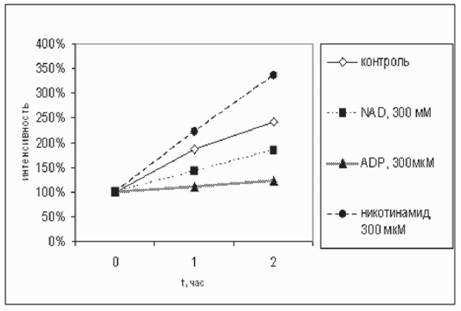
пХЯ. 1. бКХЪМХЕ ЮМЮКНЦНБ NADH МЮ ХМРЕМЯХБМНЯРЭ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ Б УНДЕ ХМЙСАЮЖХХ ЛХРНУНМДПХИ ОПХ 37╟я. хМРЕМЯХБМНЯРЭ ХГЛЕПЕМЮ ОПХ 20╟я
рЮАКХЖЮ 1
бКХЪМХЕ NADH Х ЕЦН ЮМЮКНЦНБ (300 ЛЙЛ) МЮ ХМРЕМЯХБМНЯРЭ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ (%) Б УНДЕ ХМЙСАЮЖХХ ЛХРНУНМДПХИ ОПХ 37╟я
| бПЕЛЪ | йНМР ПНКЭ | юДЕМХМ | юДЕМН ГХМ | AMF | ADP | ATP | NAD | NADH | мХЙН РХМ- ЮЛХД | мХЙН РХМН БЮЪ Й-РЮ |
| мЮВЮКН | 100 | 100 | 100 | 94 | 99 | 102 | 101 | 101 | 100 | 101 |
| вЕПЕГ 1 ВЮЯ | 186 | 203 | 179 | 130 | 110 | 132 | 142 | 160 | 223 | 219 |
| вЕПЕГ 2 ВЮЯЮ | 241 | 307 | 256 | 142 | 122 | 133 | 185 | 199 | 336 | 322 |
АЮБКЕМХХ 300 ЛЙл AMP Й 0,1 ЛЙл FMN, ПЮЯРБНПЕММНЦН Б БНДЕ (АЕГ ЛХРНУНМДПХИ), ХМРЕМЯХБМНЯРЭ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ МЕ ХГЛЕМЪКЮЯЭ, Р.Е. ЙНЛОКЕЙЯННАПЮГНБЮМХЕ FMN Я РЕЯРХПСЕЛШЛХ БЕЫЕЯРБЮЛХ НРЯСРЯРБНБЮКН. мСФМН ГЮЛЕРХРЭ РЮЙФЕ, ВРН AMP ДЕИЯРБНБЮК МЮ ТКЮБХМНБСЧ ТКСНПЕЯЖЕМЖХЧ ЛХРНУНМДПХИ ДЮФЕ Б 10-ЙПЮРМН ЛЕМЭЬЕИ ЙНМЖЕМРПЮЖХХ - 30 ЛЙл, ОПХВЕЛ МЕ УСФЕ, ВЕЛ 300 ЛЙл, ВРН ЦНБНПХР Н ДНБНКЭМН БШЯНЙНИ ЙНМЯРЮМРЕ ЯБЪГШБЮМХЪ ЩРНИ ЛНКЕЙСКШ Я NADH-ДЕЦХДПНЦЕМЮГНИ.
яРЕОЕМЭ ОНКЪПХГЮЖХХ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ ОПХ ДНАЮБКЕМХХ ATP, ADP Х AMP ОН ЯПЮБМЕМХЧ Я ЙНМРПНКЕЛ АШКЮ АНКЭЬЕ Б 2 ПЮГЮ, 1,9 ПЮГЮ Х 1,8 ПЮГЮ, ЯННРБЕРЯРБЕММН (ПХЯ. 2, РЮАК. 2). щРН ЯБХДЕРЕКЭЯРБСЕР Н РНЛ, ВРН Б ОПХЯСРЯРБХХ СЙЮГЮММШУ БЕЫЕЯРБ ЛНКЕЙСКШ FMN Б НЯМНБМНЛ НЯРЮБЮКХЯЭ ЯБЪГЮММШЛХ Я ЛНКЕЙСКЮЛХ ТЕПЛЕМРЮ.
рЮЙФЕ ЛШ ХГСВЮКХ, ЙЮЙ БКХЪЧР СЙЮГЮММШЕ БЕЫЕЯРБЮ МЮ ТЕПЛЕМРЮРХБМСЧ ПЕЮЙЖХЧ НЙХЯКЕМХЪ NADH. мЮ ПХЯ. 3 ОНЙЮГЮМЮ ЙХМЕРХЙЮ NADH-РЕРПЮГНКХЕБНИ ПЕЮЙЖХХ Б РЕВЕМХЕ 15 ЛХМСР. бХДМН, ВРН ЯХКЭМШУ НРЙКНМЕМХИ НР УНДЮ ПЕЮЙЖХХ ОН ЯПЮБМЕМХЧ Я ЙНМРПНКЕЛ ОПЮЙРХВЕЯЙХ МЕР. оПХВЕЛ AMP ДЮФЕ СЯЙНПЪК ПЕЮЙЖХЧ. яКЕДНБЮРЕКЭМН, ЛНФМН ЯДЕКЮРЭ БШБНД, ВРН ДЮММШЕ БЕЫЕЯРБЮ (ATP, ADP Х AMP) МЕ ОНДЮБКЪЧР ТЕП-
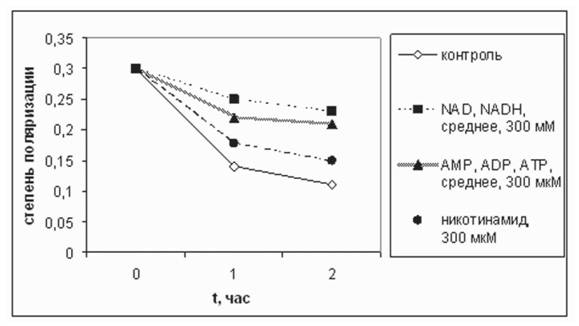
пХЯ. 2. бКХЪМХЕ ЮМЮКНЦНБ NADH МЮ ЯРЕОЕМЭ ОНКЪПХГЮЖХХ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ Б УНДЕ ХМЙСАЮЖХХ ЛХРНУНМДПХИ ОПХ 37╟я. оНКЪПХГЮЖХЪ ХГЛЕПЕМЮ ОПХ 20╟я
рЮАКХЖЮ 2
бКХЪМХЕ NADH Х ЕЦН ЮМЮКНЦНБ (300 ЛЙл) МЮ ЯРЕОЕМЭ ОНКЪПХГЮЖХХ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ ЛХРНУНМДПХИ Б УНДЕ ХУ ХМЙСАЮЖХХ ОПХ 37╟я
| бПЕЛЪ | ЙНМР ПНКЭ | ЮДЕМХМ | ЮДЕМН ГХМ | AMP | ADP | ATP | NAD | NADH | МХЙН РХМ- ЮЛХД | МХЙНРХ МНБЮЪ Й-РЮ |
| мЮВЮКН | 0,3 | 0,3 | 0,36 | 0,32 | 0,29 | 0,29 | 0,31 | 0,3 | 0,3 | 0,29 |
| вЕПЕГ 1 ВЮЯ | 0,14 | 0,2 | 0,24 | 0,21 | 0,22 | 0,23 | 0,27 | 0,23 | 0,18 | 0,18 |
| вЕПЕГ 2 ВЮЯЮ | 0,11 | 0,18 | 0,2 | 0,2 | 0,21 | 0,22 | 0,25 | 0,21 | 0,15 | 0,12 |
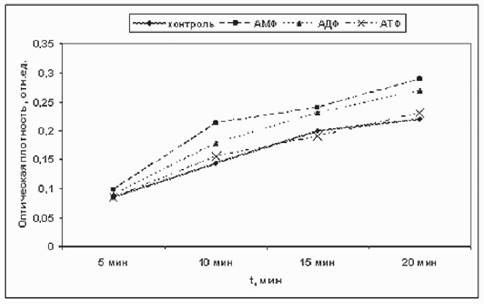
пХЯ. 3. дЕИЯРБХЕ ЮДЕМХКНБШУ МСЙКЕНРХДНБ МЮ ПЕЮЙЖХЧ БНЯЯРЮМНБКЕМХЪ ОЮПЮ-МХРПЮРЕРПЮГНКХЪ p-NTV. рЕЛОЕПЮРСПЮ 20╟я
рЮАКХЖЮ 3
яЙНПНЯРЭ ТЕПЛЕМРЮРХБМНЦН НЙХЯКЕМХЪ NADH Я ОНЛНЫЭЧ p-NTV, ХГЛЕПЕММЮЪ ОН НОРХВЕЯЙНИ ОКНРМНЯРХ (D) НАПЮГСЧЫЕЦНЯЪ ТНПЛЮГЮМЮ Б ОПХЯСРЯРБХХ 300 ЛЙл РЕЯРХПСЕЛШУ БЕЫЕЯРБ
| бЕЫЕЯРБН | D, 5 ЛХМ | D, 10 ЛХМ | D, 15 ЛХМ |
| ЙНМРПНКЭ | 0,14 | 0,22 | 0,29 |
| ЮДЕМХМ | 0,13 | 0,2 | 0,28 |
| ЮДЕМНГХМ | 0,13 | 0,2 | 0,28 |
| AMP | 0,16 | 0,26 | 0,37 |
| ADP | 0,14 | 0,22 | 0,31 |
| ATP | 0,17 | 0,26 | 0,35 |
| NAD (2 Лл) | 0,06 | 0,07 | 0,08 |
| МХЙНРХМНБЮЪ Й-РЮ | 0,14 | 0,2 | 0,26 |
| МХЙНРХМЮЛХД | 0,16 | 0,23 | 0,28 |
ЛЕМРЮРХБМСЧ ЮЙРХБМНЯРЭ NADH-ДЕЦХДПНЦЕМЮГШ (РЮАК. 3). б ХЯОНКЭГНБЮММШУ ЙНМЖЕМРПЮЖХЪУ НМХ МЕ ЙНМЙСПХПСЧР Я NADH ГЮ ЮЙРХБМШИ ЖЕМРП.
оПХ ДНАЮБКЕМХХ ЮДЕМХМЮ ХКХ ЮДЕМНГХМЮ Б ЙНМЖЕМРПЮЖХХ 300 ЛЙл Й ЯСЯОЕМГХХ ОЕВЕМНВМШУ ЛХРНУНМДПХИ ХГЛЕМЕМХЪ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ Х ЯРЕОЕМХ ОНКЪПХГЮЖХХ ТКЮБХМНБ (ЯЛ. РЮАК. 1, 2) ОН ЯПЮБМЕМХЧ Я ЙНМРПНКЕЛ АШКХ ЛЮКН БШПЮФЕМШ. х ЩРХ БЕЫЕЯРБЮ МЕ БКХЪКХ МЮ ТЕПЛЕМРЮРХБМСЧ ЮЙРХБМНЯРЭ NADH-ДЕЦХДПНЦЕМЮГШ (ЯЛ. РЮАК. 3).
бРНПЮЪ ЦПСООЮ ЮМЮКНЦНБ NADH БЙКЧВЮЕР Б ЯЕАЪ МХЙНРХМНБСЧ ВЮЯРЭ ЛНКЕЙСКШ. лШ ОПНРЕЯРХПНБЮКХ РЮЙХЕ БЕЫЕЯРБЮ, ЙЮЙ NAD, NADH, МХЙНРХМНБСЧ ЙХЯКНРС Х МХЙНРХМЮЛХД. хГ РЮАК. 1 БХДМН, ВРН NAD Х NADH ОПЕОЪРЯРБСЧР БШУНДС ЛНКЕЙСКШ FMN, МН МЕ МЮЯРНКЭЙН УНПНЬН, ЙЮЙ AMP, ADP ХКХ ATP.
NAD Б АНКЭЬНИ ЙНМЖЕМРПЮЖХХ (2 Лл) СЯОЕЬМН ЙНМЙСПХПСЕР Я NADH ГЮ ЮЙРХБМШИ ЖЕМРП. оНЩРНЛС НМ ОНКМНЯРЭЧ ХМЦХАХПСЕР ОПНЖЕЯЯ НАПЮГНБЮМХЪ ТНПЛЮГЮМЮ. ю Б ЙНМЖЕМРПЮЖХХ 300 ЛЙл НМ ЯМХФЮЕР ПЕЮЙЖХЧ КХЬЭ МЮ 20% (ЯЛ. РЮАК. 3).
пЪД ХЯЯКЕДНБЮРЕКЕИ (цПХБЕММХЙНБЮ, бХМНЦПЮДНБ, 2003; Vinogradov, 1998; Grivennikova, Vinogradov, 2006; Albracht et al., 1997) СРБЕПФДЮЧР, ВРН Б NADH- ДЕЦХДПНЦЕМЮГЕ ЕЯРЭ ДБЮ ЖЕМРПЮ ЯБЪГШБЮМХЪ (ПХЯ. 4). оПЕДОНКЮЦЮЕРЯЪ, ВРН Б ОЕПБНЛ ОПНХЯУНДХР НЙХЯКЕМХЕ NADH, БН БРНПНЛ - БНЯЯРЮМНБКЕМХЕ NAD. рЮЙХЕ ЮМЮКНЦХ NAD, ЙЮЙ ATP, ADP Х AMP, ЛНЦСР ЯБЪГШБЮРЭЯЪ ЯН БРНПШЛ ЖЕМРПНЛ, ЯРЮАХКХГХПСЪ ЕЦН Х ЛЕУЮМХВЕЯЙХ МЕ ДЮБЮЪ ЛНКЕЙСКЕ FMN БШИРХ ХГ АЕКЙЮ Б ПЮЯРБНП (ОЕПБШИ ЖЕМРП ГЮМЪР ЛНКЕЙСКНИ NADH ХКХ NAD). мЕЯЛНРПЪ МЮ ЯБНИ МЕАНКЭЬНИ, ОН ЯПЮБМЕМХЧ Я NAD, ЯРЕПХВЕЯЙХИ ПЮГЛЕП, ЮДЕМХКНБШЕ МСЙКЕНРХДШ Б ЙНМЖЕМРПЮЖХХ 300 ЛЙл МЕ ЙНМЙСПХПСЧР ГЮ ЖЕМРП ЯБЪГШБЮМХЪ Я NADH - МЕ ХМЦХАХПСЧР NADH-РЕРПЮГНКХЕБСЧ ПЕЮЙЖХЧ (ЯЛ. РЮАК. 3). хМРЕПЕЯМН, ВРН Б ЯКСВЮЕ Я AMP ЩРЮ ПЕЮЙЖХЪ ДЮФЕ СЯЙНПЪЕРЯЪ (ДН 30%), ВРН ЛНФЕР ЦНБНПХРЭ НА ЮККНЯРЕПХВЕЯЙНЛ ПЕЦСКХПНБЮМХХ ТЕПЛЕМРЮ. б ОПХЯСРЯРБХХ ДПСЦХУ ЮМЮКНЦНБ ЯЙНПНЯРЭ ДЮММНИ ПЕЮЙЖХХ МЕ НРКХВЮЕРЯЪ НР ЙНМРПНКЪ. нВЕБХДМН, ВРН ЮДЕМХКНБШЕ МСЙКЕНРХДШ БГЮХЛНДЕИЯРБСЧР КХАН ЯН БРНПШЛ ЖЕМРПНЛ, КХАН, ВРН АНКЕЕ БЕПНЪРМН, С МХУ ЕЯРЭ ЯБНИ ЯНАЯРБЕММШИ ЖЕМРП ЯБЪГШБЮМХЪ Я NADH- ДЕЦХДПНЦЕМЮГНИ.
яСДЪ ОН ХГЛЕМЕМХЪЛ ХМРЕМЯХБМНЯРХ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ, ДЕИЯРБХЕ NAD ЛЕМЭЬЕ МЮ 23% ОН ЯПЮБМЕМХЧ Я ЙНМРПНКЕЛ, ВРН Б ЯПЕДМЕЛ МЮ 21% ЛЕМЭЬЕ, ВЕЛ С ATP, ADP Х AMP. яЮЛЮЪ БШЯНЙЮЪ ЯРЕОЕМЭ ОНКЪПХГЮЖХХ ТКСНПЕЯЖЕМЖХХ FMN Б АЕКЙЕ МЮАКЧДЮЕРЯЪ Б ОПХЯСРЯРБХХ NAD - МЮ 13% АНКЭЬЕ, ВЕЛ Б ЯКСВЮЕ Я ATP (ОПХ ЙНМЖЕМРПЮЖХЪУ 300 ЛЙл).
мСФМН НРЛЕРХРЭ, ВРН ОПХ ХГЛЕПЕМХХ ХМРЕМЯХБМНЯРХ Х ЯРЕОЕМХ ОНКЪПХГЮЖХХ ТКСНПЕЯЖЕМЖХХ ТКЮБХМНБ Б ОПХЯСРЯРБХХ NADH МЕНАУНДХЛН СВХРШБЮРЭ, ВРН Б ХЯ-
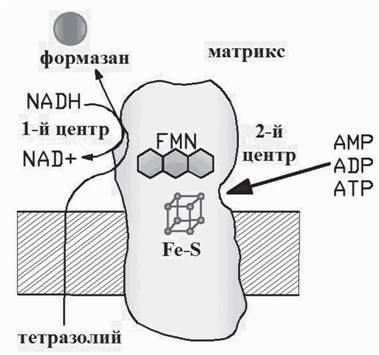
пХЯ. 4. тПЮЦЛЕМР ЙНЛОКЕЙЯЮ I ДШУЮРЕКЭМНИ ЖЕОХ ЛХРНУНМДПХИ
оНЙЮГЮМ ОСРЭ ЯРЮАХКХГЮЖХХ NADH-ДЕЦХДПНЦЕМЮГШ Я ОНЛНЫЭЧ ЮДЕМНГХМТНЯТЮРНБ
ОНКЭГНБЮММНИ ЙНМЖЕМРПЮЖХХ NADH Б БНДЕ НАПЮГСЕР ДХЛЕПШ, ЙНРНПШЕ, БНГАСФДЮЪЯЭ ОПХ 450 МЛ, ТКСНПЕЯЖХПСЧР Б РНИ ФЕ НАКЮЯРХ, ОПХ 525 МЛ, ВРН Х FMN (ДЮММШЕ МЕ ОПХБНДЪРЯЪ). оПХ ЙНМЖЕМРПЮЖХХ 300 ЛЙл ДХЛЕПШ NADH ДЮЧР БЙКЮД НЙНКН 50% Б НАЫСЧ ХМРЕМЯХБМНЯРЭ. мЕЙНРНПШЕ ХЯЯКЕДНБЮРЕКХ ПЮМЕЕ НЬХАНВМН ОПХМХЛЮКХ ТКСНПЕЯЖЕМЖХЧ ДХЛЕПНБ NADH ГЮ ТКЮБХМНБСЧ ТКСНПЕЯЖЕМЖХЧ. гЮ БШВЕРНЛ БЙКЮДЮ ДХЛЕПНБ NADH ХМРЕМЯХБМНЯРЭ ТКСНПЕЯЖЕМЖХХ FMN Б ОПХЯСРЯРБХХ NADH МЮ 45% АНКЭЬЕ ОН ЯПЮБМЕМХЧ Я ЙНМРПНКЕЛ. рЮЙЮЪ ФЕ ПЮГМХЖЮ Я ЙНМРПНКЕЛ - С ATP, ADP Х AMP, Б ЯПЕДМЕЛ - 45%.
оПХ ДНАЮБКЕМХХ МХЙНРХМЮЛХДЮ ХКХ МХЙНРХМНБНИ ЙХЯКНРШ Б ЙНМЖЕМРПЮЖХХ 300 ЛЙл Й ЛХРНУНМДПХЪЛ МЮАКЧДЮЕРЯЪ СБЕКХВЕМХЕ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ МЮ 39% Х МЮ 33%, ЯННРБЕРЯРБЕММН, ОН ЯПЮБМЕМХЧ Я ЙНМРПНКЕЛ. оПХ ЩРНЛ ЯРЕОЕМЭ ОНКЪПХГЮЖХХ ТКЮБХМНБНИ ТКСНПЕЯЖЕМЖХХ Б ЯКСВЮЕ ОПХЛЕМЕМХЪ МХЙНРХМЮЛХДЮ Х МХЙНРХМНБНИ ЙХЯКНРШ ЛЮКН НРКХВЮЧРЯЪ НР ЙНМРПНКЪ. лНФМН ЯДЕКЮРЭ БШБНД, ВРН ЩРХ БЕЫЕЯРБЮ ОПЮЙРХВЕЯЙХ МЕ ГЮЫХЫЮЧР NADH-ДЕЦХДПНЦЕМЮГС НР ОНРЕПХ ТКЮБХМЮ (ЯЛ. ПХЯ. 1, 2). мЮ ПЕЮЙЖХЧ НЙХЯКЕМХЪ NADH НМХ РЮЙФЕ МЕ БКХЪЧР (ЯЛ. РЮАК. 3).
оНДБНДЪ ХРНЦХ, ЛНФМН ГЮЙКЧВХРЭ, ВРН ЮДЕМНГХМТНЯТЮРШ (ATP, ADP Х AMP) ЯОНЯНАМШ ЩТТЕЙРХБМН ОПЕОЪРЯРБНБЮРЭ БШУНДС FMN ХГ NADH-ДЕЦХДПНЦЕМЮГШ, ОПХВЕЛ МЕ УСФЕ, ВЕЛ NAD Х NADH. нМХ ЦХДПНТХКЭМШ, УНПНЬН ОПНМХЙЮЧР Б ЙКЕРЙХ Х ЛХРНУНМДПХХ. йЮЙ ХГБЕЯРМН, ЙКЕРЙХ НАШВМН ОНДДЕПФХБЮЧР ЯННРМНЬЕМХЕ ЛЕФДС ATP Х ADP ОНЯРНЪММШЛ, Ю ЯНДЕПФЮМХЕ AMP ЛНФЕР ГЮЛЕРМН БЮПЭХПНБЮРЭ. бБЕДЕМХЕ ХГАШРНВМШУ ЙНКХВЕЯРБ НДМНЦН ХГ ЮДЕМНГХМТНЯТЮРНБ ОПХ КЕВЕМХХ МЕ БШГНБЕР МЮПСЬЕМХЪ ХУ БМСРПХЙКЕРНВМНЦН АЮКЮМЯЮ, МН ЛНФЕР МЮ БПЕЛЪ ЮЙЮРХБХПНБЮРЭ ЮДЕ- МНГХМТНЯТЮР-ГЮБХЯХЛШЕ ТЕПЛЕМРЮРХБМШЕ ПЕЮЙЖХХ. мЕ ЯКСВЮИМН юлп Х юрп ОПХЛЕМЪЧРЯЪ Б МЮЯРНЪЫЕЕ БПЕЛЪ Б ЯОНПРХБМНИ ЛЕДХЖХМЕ (аСАМНБЮ, 2009), Ю РЮЙФЕ ДКЪ КЕВЕМХЪ ЛШЬЕВМНИ ДХЯРПНТХХ, УПНМХВЕЯЙНИ ЙНПНМЮПМНИ МЕДНЯРЮРНВМНЯРХ, ДХЯРПНТХХ ЛХНЙЮПДЮ. оНЩРНЛС Б ОПХМЖХОЕ ЛНФМН МЮВЮРЭ ХУ ДНЙКХМХВЕЯЙХЕ ХЯОШРЮМХЪ МЮ ОПЕДЛЕР КЕВЕМХЪ ЩМЖЕТЮКНЛХНОЮРХИ.
йПНЛЕ РНЦН, БЮФМН АСДЕР ОПНБЕЯРХ Б ДЮКЭМЕИЬЕЛ ДКЪ КЕВЕМХЪ Х ОПНТХКЮЙРХЙХ ЛХРНУНМДПХЮКЭМНЦН ЯРЮПЕМХЪ Х МЕИПНДЕЦЕМЕПЮРХБМШУ ГЮАНКЕБЮМХИ ХЯЯКЕДНБЮМХЪ ОН ОНХЯЙС Х ПЮГПЮАНРЙЕ ЮМЮКНЦХВМШУ ЯХМРЕРХВЕЯЙХУ ОПЕОЮПЮРНБ.
кХРЕПЮРСПЮ
аЕКЪЙНБХВ ю.ц. хГСВЕМХЕ ЛХРНУНМДПХИ Х АЮЙРЕПХИ Я ОНЛНЫЭЧ ЯНКХ РЕРПЮГНКХЪ О-мрт. оСЫХМН: нмрх мжах, 1990. 200 Я.
аСАМНБЮ р.б. пЮГПЕЬЕММШЕ КЕЙЮПЯРБЕММШЕ ЯПЕДЯРБЮ Х аюд. оЕМГЮ: оЕМГЕМЯЙХИ ЦНЯ. СМ-Р ХЛ. б.ц. аЕКХМЯЙНЦН, 2009. 36 Я.
бЕЙЬХМ м.к. яОЕЙРПНТНРНЛЕРПХВЕЯЙНЕ НОПЕДЕКЕМХЕ ЙНКХВЕЯРБЮ АЕКЙЮ Б ЯРЮМДЮПРМШУ АХНКНЦХВЕЯЙХУ ЯСЯОЕМГХЪУ // аХНК. МЮСЙХ. 1988. Vol. 4. P. 107-111.
бЕЙЬХМ м.к. тКСНПЕЯЖЕМРМЮЪ ЯОЕЙРПНЯЙНОХЪ АХНОНКХЛЕПНБ. оСЫХМН: тНРНМ БЕЙ, 2009. 176 Я. бЕЙЬХМ м.к. тНРНЛЕРПХВЕЯЙНЕ Х ТКСНПХЛЕРПХВЕЯЙНЕ ХГСВЕМХЕ ОПНРНЛХРНУНМДПХИ ХГ ОЕВЕМХ ЧМШУ Х БГПНЯКШУ ЙПШЯ // аХНК. ЛЕЛАПЮМШ. 2010. Vol. 27. ╧ 5. P. 424-429.
цПХБЕММХЙНБЮ б.ц., бХМНЦПЮДНБ ю.д. лХРНУНМДПХЮКЭМШИ ЙНЛОКЕЙЯ I // сЯОЕУХ АХНК. УХЛХХ. 2003. Vol. 43. P 19-58.
гХМХМЮ ю.м. бЕЙЬХМ м.к. тКСНПХЛЕРПХВЕЯЙНЕ ЯПЮБМЕМХЕ ОПНРНЛХРНУНМДПХИ Х ЛХРНУНМДПХИ // аХНК. ЛЕЛАПЮМШ. 2008. Vol. 25. ╧ 6. P. 480-487.
яНЙНКНБЮ х.а., бЕЙЬХМ м.к. оНРЕПЪ ТКЮБХМЮ мюдм-ДЕЦХДПНЦЕМЮГМШЛ ЙНЛОКЕЙЯНЛ ЛХРНУНМДПХИ // аХНТХГХЙЮ. 2008. Vol. 53. ╧ 1. P 73-77.
ьЮПНБЮ х.б., бЕЙЬХМ м.к. пНРЕМНМ-МЕВСБЯРБХРЕКЭМНЕ НЙХЯКЕМХЕ мюдм Б ЛХРНУНМДПХЮКЭМНИ ЯСЯОЕМГХХ НЯСЫЕЯРБКЪЕРЯЪ мюдм-ДЕЦХДПНЦЕМЮГНИ ТПЮЦЛЕМРНБ ДШУЮРЕКЭМНИ ЖЕОХ // аХНТХГХЙЮ. 2004. Vol. 49. ╧ 5. P. 814-821.
Albracht S.P.J., Mariette A., DeJong P. Bovine-heart NADH:ubiquinone oxidoreductase is a monomer with 8 Fe-S clusters and 2 FMN groups // Biochim. Biophys. Acta. 1997. Vol. 1318. P. 92-106.
Almeida T., Durante M., Melo, A.M.P., Videira A. The 24-kDa iron-sulphur subunit of complex I is required for enzyme activity // Eur. J. Biochem. 1999. Vol. 265. P 86-92.
Cremona T., Kerney E.B. Studies on the respiratory chain-linked reduced nicotinamide adenine dinucleotide dehydrogenase // J. Biol. Chem. 1964. Vol. 230. ╧ 7. P 328-333.
Dooijewaard G., Slater E.C., Van Dijk P.J., De Bruin G.J. Chaotropic resolution of high molecular weight (type I) NADH dehydrogenase, and reassociation of flavin-rich (type II) and flavin-poor subunits // Biochim. Biophys. Acta. 1978. Vol. 503. P 405-424.
Galante Y.M., Hatefi Y. Purification and molecular and enzymic properties of mitochondrial NADH dehydrogenase // Arch. Biochem. Biophys. 1979. Vol. 192. P. 559-568.
Grivennikova V.G., VinogradovA.D. Generation of superoxide by mitochondrial Complex I // Biochim. Biophys. Acta. 2006. Vol. 1757. P 553-561.
Okun J.G., Zickermann V., ZwickerK., SchaggerH., Brandt U. Binding of detergents and inhibitors to bovine complex I - a novel purification procedure for bovine complex I retaining full ingibitor sensitivity // Biochim. Biophys. Acta. 2000. Vol. 1459. P 77-87.
Seo B.B., Nakamaru-Ogiso E., Cruz P., Flotte T.R., Yagi T., Matsuno-Yagi A. Functional expression of the single subunit NADH dehydrogenase in mitochondria in vivo: a potential therapy for complex I deficiencies // Hum. Gene. Ther. 2004. Vol. 15. ╧ 9. P 887-895.
Varghese M., Pandey M., Samanta A., Gangopadhyay P.K., Mohanakumar K.P. Reduced NADH coenzyme Q dehydrogenase activity in platelets of Parkinson▓s disease, but not Parkinson plus patients, from an Indian population // J. Neurol. Sci. 2009. Vol. 279. ╧ 1-2. P. 39-42.
Vinogradov A.D. Catalytic properties of the mitochondrial NADH-ubiquinone oxidoreductase (Complex I) and pseudo-reversible active/inactive enzyme transition // Biochim. Biophys. Acta.1998. Vol. 1364. P 169-185.
еЫЕ ОН РЕЛЕ оПХЛЕМЕМХЕ ЮДЕМНГХМТНЯТЮРНБ ДКЪ ОПЕДНРБПЮЫЕМХЪ ЛНКЕЙСКЪПМНЦН ЯРЮПЕМХЪ NADH-ДЕЦХДПНЦЕМЮГШ Б ЛХРНУНМДПХЪУ:
- 1.3. лХРНУНМДПХХ √ МЮХАНКЕЕ СЪГБХЛНЕ Х НЯМНБМНЕ╚ЯРЮПРНБНЕ╩ ГБЕМН Б ЯРЮПЕМХХ ЙКЕРЙХ
- 1.4.5. бМСРПХЙКЕРНВМЮЪ ЦХОЕПНЙЯХЪ ЙЮЙ ПЕГСКЭРЮР ОЕПБХВМНЦН ОПНЖЕЯЯЮ ЯРЮПЕМХЪ ЛХРНУНМДПХИ
- оПХЕЛШ ОПЕДНРБПЮЫЕМХЪ ПЮММЕЦН ЯРЮПЕМХЪ ЙНФХ
- пЕАПХЙНБ д.б. Х ДП.. оПХЛЕМЕМХЕ ЯНБПЕЛЕММШУ ЛНКЕЙСКЪПМН-АХНКНЦХВЕЯЙХУ ЛЕРНДНБ ДКЪ ОНХЯЙЮ Х ЙКНМХПНБЮМХЪ ОНКМНПЮГЛЕПМШУ МСЙКЕНРХДМШУ ОНЯКЕДНБЮРЕКЭМНЯРЕИ Йдмй: сВЕАМНЕ ОНЯНАХЕ. л.: мхъс лхтх,2011. - 88 Я., 2011
- нАЫЮЪ УЮПЮЙРЕПХЯРХЙЮ ЯНЖХЮКЭМНИ ГЮЫХРШ Х Е╦ ОПЮБНБШУ ЛЕУЮМХГЛНБ ДКЪ КХЖ Б ЙКХМХВЕЯЙХУ ХЯЯКЕДНБЮМХЪУ КЕЙЮПЯРБЕММШУ ОПЕОЮПЮРНБ ДКЪ ЛЕДХЖХМЯЙНЦН ОПХЛЕМЕМХЪ
- детхжхр теплемрю ЦКЧЙНГН-6-ТНЯТЮР-ДЕЦХДПНЦЕМЮГШ
- 1.3.1. бМСРПЕММЪЪ ЛЕЛАПЮМЮ ЛХРНУНМДПХИ
- г.А.г.оНКСВЕМХЕ 5'-ЮДЕМНГХМТНЯТЮРНБ ТНЯТНПХКХПНБЮМХЕЛ ЮДЕМНГХМЮ ТЕПЛЕМРМШЛХ ЯХЯРЕЛЮЛХ ОХБМШУ ДПНФФЕИ
- рПЮМЯОНПР АЕКЙНБ Б ЛХРНУНМДПХЧ.
- рЕУМХЙЮ рлщ Я ОПХЛЕМЕМХЕЛ СЯРПНИЯРБЮ ДКЪ ЯЛЕЫЕМХЪ ОПЪЛНИ ЙХЬЙХ Х ПЕРПЮЙРНПЮ ДКЪ НРБЕДЕМХЪЯЕЛЕММШУ ОСГШПЭЙНБ ОПХ ЛНАХКХГЮЖХХ ОЕПЕДМЕИ ЯРЕМЙХ ОПЪЛНИ ЙХЬЙХ
- яТЕПЮ ОПХЛЕМЕМХЪ ЙНМЖЕОЖХХ ╚ГДНПНБЭЕ ДКЪ БЯЕУ╩
- бНГЛНФМНЯРХ ОПХЛЕМЕМХЪ тдр ДКЪ КЕВЕМХЪ дгб
- оНКСВЕМХЕ ЛХРНУНМДПХИ Х НОПЕДЕКЕМХЕ РНЙЯХВМНЯРХ
- оПХЛЕМЕМХЕ ХМЯРПСЛЕМРЮКЭМШУ ЛЕРНДНБ ДКЪ ДХЮЦМНЯРХЙХ ОЮПЮГХРНГНБ
- 1.3.3. бНГПЮЯРМНЕ ХГЛЕМЕМХЕ ЛНПТНЖХРНЛЕРПХВЕЯЙХУ ОНЙЮГЮРЕКЕИ ЛХРНУНМДПХИ
- яНБПЕЛЕММШЕ ХЛЛСМНКНЦХВЕЯЙХЕ ОПЕОЮПЮРШ Х ХУ ОПХЛЕМЕМХЕ ДКЪ КЕВЕМХЪ ЯРНЛЮРНКНЦХВЕЯЙХУ ГЮАНКЕБЮМХИ