Заключение
Известно, что такие медиаторы иммунной системы, как антивоспалительные цитокины, продуцируются резидентными клетками мозга, прежде всего микроглией и астроцитами, при нормальных физиологических условиях (Szelenyl, 2001).
Однако при повреждениях мозга, связанных с нейрологическими заболеваниями, механической травмой, гипоксией/ишемией или инфекциями, экспрессия антивоспалительных цитокинов резко возрастает. Современные экспериментальные и клинические наблюдения указывают на их протектирующее действие в отношении повреждения нейронов в ответ на стрессовые (потенциально летальные) стимулы (Strle et al., 2001; Zhou et al, 2009; DeBilbao et al., 2009). Связывание антивоспалительного цитокина ИЛ-10 с его мембранным рецептором приводит к активации нескольких канонических внутриклеточных сигнальных путей, включающих: Jak/ Stat, MEK/ERK и PI3 киназа/Akt (Strle et al., 2001). Экспериментальные данные показывают, что ИЛ-10 через стимуляцию Jak/Stat сигнального пути способен регулировать активность PI3 киназа/Akt, MEK/ERK и фактор транскрипции NF- kB, что, в свою очередь, приводит к активации генов, продукты которых, с одной стороны, ингибируют апоптоз, а с другой - стимулируют протектирующие механизмы клеток организма (Strle et al., 2001; Leker, Shohami, 2002). Модулирующие эффекты ИЛ-10, опосредованные этой цепочкой событий, развиваются с большой задержкой и требуют для своей реализации несколько часов - суток. Однако эти экспериментальные данные оказались неполными. Относительно недавно было показано, что в центральной нервной системе позвоночных такие сигнальные молекулы, как цитокины, наряду с их ролью в регуляции иммунного ответа и воспаления способны также модулировать возбудимость нервных клеток через лиганд- и потенциал-зависимые мембранные каналы, синаптическую передачу и нейрональную пластичность. Причем, в отличие от классических представлений о действии цитокинов, влияние этих медиаторов иммунной системы на активность нейронов может наблюдаться уже в течение нескольких минут после их приложения к мембране нервных клеток (быстрые эффекты) (Beattie et al., 2002; Viviani et al., 2003; Stellwagen et al., 2005; Schaffers, Sorkin, 2008; Turovskaya et al., 2012; Savina et al., 2013; Turovsky et al., 2013).Результаты проведенных нами исследований позволили получить принципиально новые данные о роли кальций-зависимых калиевых каналов, фоновых 2Р калиевых каналов (предварительные результаты), внутриклеточных депо ионов кальция, Ca2+/кальмодулин-зависимой протеинкиназы II и ионотропных рецепторов глутамата в механизмах быстрого модулирующего действия антивоспалительного цитокина ИЛ-10, через которые данный цитокин способен оказывать нейропротек- тирующие действия на повреждения клеток мозга, индуцируемых гипоксией или ишемией.
На рисунке представлена принципиальная схема пирамидного нейрона поля СА1 гиппокампа с клеточными мишенями (показаны крупными стрелками) модулирующего действия ИЛ-10 по результатам наших исследований. Гипоксия или ишемия способны инициировать три группы событий в этих нейронах: уменьшать вероятность генерации потенциалов действия, инициировать трансформацию одиночного потенциала действия в пачки потенциалов действия и приводить к гибели этих нейронов. Антивоспалительный цитокин ИЛ-10, модулируя соответствующие клеточные мишени, оказывает нейропротектирующее действие на индуцируемые гипоксией или ишемией нарушения в их активности.
Современные клинические данные и результаты экспериментальных исследований на животных показывают, что центральная роль в развитии нейродегенеративных процессов при ишемии/гипоксии, механической травме, эпилепсиях, а также при таких нейродегенеративных заболеваниях мозга, как болезнь Альцгеймера или болезнь Паркинсона, принадлежит провоспалительным цитокинам. В связи с этим одним из перспективных подходов для разработки нового класса нейропротекторов может быть использование эндогенных антивоспалительных цитокинов. Из фундаментальных исследований также известно, что опосредованная глутаматом эксайтотоксичность, а также нарушение гомеостаза ионов кальция являются одними из ключевых детерминантов клеточной гибели при этих нейро-
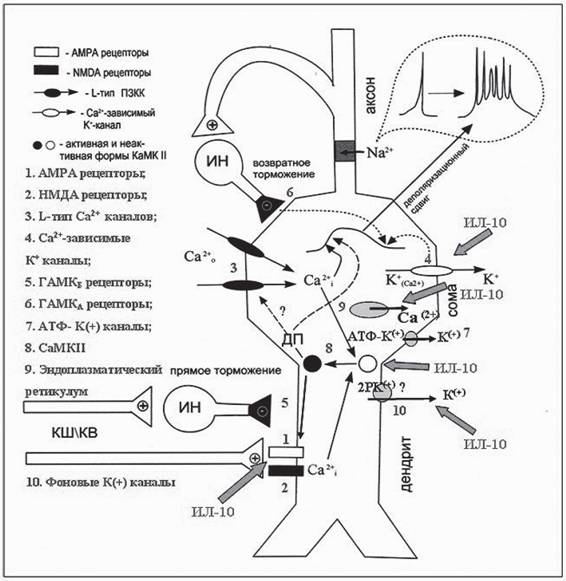
Рис.
Принципиальная схема пирамидного нейрона поля СА1 гиппокампа с клеточными мишенями (показаны крупными стрелками) модулирующего действия ИЛ-10 по результатам проведенных нами исследований1 - АМРА рецепторы; 2 - NMDA рецепторы; 3 - L-тип Са2+ каналов; 4 - Са2+-зависимые К+-кана- лы; 5 - ГАМКб рецепторы; 6 - ГАМКа рецепторы; 7 - АТР-К+ -АТФ-зависимые К+ каналы; 8 - КаМК II - Са2+/кальмодулин-зависимая протеинкиназа II; 9 - Инозитолтрифосфат-зависимые участки эндоплазматического ретикулума; 10 - фоновые 2Р калиевые каналы. ДП - долговременная потенциация
патологиях. Результаты представленных нами исследований позволили показать на разных моделях ишемии in vivo и in vitro, а также на in vitro моделях гипоксии, что антивоспалительный цитокин ИЛ-10 оказывает нейропротектирующее действие на повреждение нейронов мозга, индуцируемых ишемией или гипоксией. Кроме того, полученные результаты позволили получить принципиально-новые представления о роли различных мембранных калиевых каналов, внутриклеточных депо ионов кальция, Са2+/кальмодулин-зависимой протеинкиназы II и ионотропных рецепторов глутамата в механизмах нейропротектирующего действия ИЛ-10 на повреждения нейронов мозга, индуцируемых гипоксией или ишемией. Тем не менее, необходимо продолжить исследования по анализу механизмов этого действия антивоспалительных цитокинов. Одним из перспективных подходов для разработки нового класса нейропротекторов может быть использование активаторов стимулируемых ИЛ-10 протектирующих внутриклеточных сигнальных каскадов, способных блокировать сигнальные каскады гибели клеток мозга. Из литературных данных известно, что в клетках иммунной системы активируемый ИЛ-10 PI3 киназа/ Akt внутриклеточный сигнальный каскад является одним из ключевых каскадов, опосредующих активацию генов, продукты которых, с одной стороны, ингибируют апоптоз, а с другой - стимулируют протектирующие механизмы клеток. Однако нейропротектирующая роль этого сигнального каскада в клетках мозга остается неясной.
Поэтому исследования в этом направлении могут дать новые, имеющие практическое значение для медицины, представления о роли и механизмах ней- ропротектирующего действия антивоспалительных цитокинов как на индуцируемые ишемией или гипоксией повреждения нейронов мозга, так и на патогенез таких нейродегенеративных заболеваний мозга как болезни Альцгеймера или Паркинсона и как следствие - разработать на основе этих знаний новые лекарственные препараты.Литература
Левин С.Г., Годухин О.В. Роль Са2+-активируемых калиевых каналов большой проводимости в прекондиционирующих эффектах гипоксии и постгипоксической гипервозбудимости нейронов поля СА1 гиппокампа in vitro // ЖВНД. 2009. Вып. 59. № 5. С. 622-629.
Левин С.Г., Шамсутдинова А.А., Годухин О.В. Сравнительное действие блокаторов АТФ-чувствитель- ных К+-каналов на индуцируемые кратковременными эпизодами гипоксии изменения активности пирамидных нейронов поля СА1 срезов гиппокампа // Бюлл. эксперим. биол. и медицины. 2012. Вып. 154. № 10. С. 435-438.
Левин С.Г., Конаков М.В., Годухин О.В. К механизмам действия интерлейкина-10 на индуцируемые гипоксией изменения в активности нейронов гиппокампа: возможная роль 2Р калиевых каналов. // Рецепторы и внутриклеточная сигнализация. Ред. В.П. Зинченко, А.В. Бережнов. Пущино: Ема, 2013. Т 2. С. 457-461.
Asadullan K., Sterry W., Volk H.D. Interleukin-10 therapy - review of a new approach // Pharmacol. Rev. 2003. Vol. 55. № 2. Р 241-269.
Beattie E.C., Stellwagen D., Morishita W. et al. Control of synaptic strength by glial TNFa // Science. 2002. Vol. 295. P. 2282-2285.
De Bilbao F., Arsenijevic D., Moll T. et al. In vivo over-expression of interleukin-10 increases resistance to focal brain ischemia in mice // J. Neurochem. 2009. Vol. 110. P 12-22.
Gao T.M., Fung M.L. Decreased large conductance Ca^-activated Kw channel activity in dissociated CA1 hippocampal neurons in rats exposed to perinatal and postnatal hypoxia // Neurosci. Lett.
2002. Vol. 332. № 3. P. 163-166.Harukuni I., Bhardwaj A. Mechanisms of brain injury after global cerebral ischemia // Neurol. Clin. 2006. Vol. 24. P. 1-21.
Kulik A., Brockhaus J., Pedarzani P., Ballanyi K. Chemical anoxia activates ATP-sensitive and blocks Ca2+- dependent K+ channels in rat dorsal vagal neurons in situ // Neuroscience. 2002. Vol. 110. № 3. P. 541-54.
Leker R.R., Shohami E. Cerebral ischemia and trauma - different etiologies yet similar mechanisms: neuroprotective opportunities // Brain Res. Rev. 2002. Vol. 39. P. 55-73.
Levin S.G., Godukhin O.V. Comparative roles of ATP-sensitive K+ channels and Ca2+-activated BK+ channels in posthypoxic hyperexcitability and rapid hypoxic preconditioning in hippocampal CA1 pyramidal neurons in vitro // Neurosci. Lett. 2009. Vol. 461. P. 90-94.
Levin S.G., Godukhin O.V Anti-inflammatory cytokines, TGF-P1 and IL-10, exert anti-hypoxic action and abolish posthypoxic hyperexcitability in hippocampal slice neurons: Comparative aspects // Exper. Neurology. 2011. Vol. 232. P. 329-332.
Marchetti B., Abbracchio M.P. To be or not to be (inflamed) - is that the question in anti-inflammatory drug therapy of neurodegenerative disorders? // Trends in Pharmac. Sci. 2005. Vol. 26. № 10. P. 517-525.
Martin L.J. The apoptosis - necrosis cell death continuum in the CNS development, injury and disease: contribution and mechanisms // Neuroprotection. Eds E.N. Lo, J. Marwah. USA, Prominenet Press, 2002, P. 380-412.
Mehta S.L., Manchas N., Raghubir R. Molecular targets in cerebral ischemia for developing novel therapeutics // Brain Res. Rev. 2007. Vol. 54. P. 34-66.
MichaudK., ForgetH., Cohen H. Chronic glucocorticoid hypersecretion in Cushing’s syndrome exacerbates cognitive aging // Brain and Cognition. 2009. Vol. 71. P. 1-8.
NixonR.A. The role of autophagy in neurodegenerative disease // Nature Medicine 2013. Vol. 19. P. 983-997. doi:10.1038/nm.3232.
Obrenovitch T.P. Molecular physiology and preconditioning-induced brain tolerance to ischemia // Physiol.
Rev. 2008. Vol. 88. P. 211-247.PlantL.D., Kemp P.J., Peers D.C., Henderson Z., Pearson H.A. Hypoxic depolarization of cerebellar granule neurons by specific inhibition of TASK-1 // Stroke. 2002. Vol. 33. P. 2324-2328.
Puyal J., Ginet V., Peter G.H. Clarke P.G.H. Multiple interacting cell death mechanisms in the mediation of excitotoxicity 3 and ischemic brain damage: A challenge for neuroprotection // Progress in Neurobiology. 2013. dx.doi.org/10.1016/j.pneurobio.2013.03.002.
Savina T.A., Shchipakina T.G., Levin S.G., Godukhin O.V. Interleukin-10 prevents the hypoxia-induced decreases in expressions of AMPA receptor subunit GluA1 and alpha subunit of Ca2+/calmodulin-depen- dent protein kinase II in hippocampal neurons // Neurosci. Lett. 2013. Vol. 534. P. 279-284.
Schaffers M., Sorkin L. Effect of cytokines on neuronal excitability // Neurosci. Lett. 2008. Vol. 437. P. 188-193.
Stankowski J.N., Gupta R. Therapeutic targets for neuroprotection in acute ischemic stroke: lost of translation? // Antioxidants and Redox Signaling. 2011. Vol. 14. № 10. P. 1841-1851.
Stellwagen D., Beattie E.C., Seo J.Y., Malenka R.C. Differential regulation of AMPA receptor and GABA receptor trafficking by tumor necrosis factor-а // J. Neurosci. 2005. Vol. 25. № 12. P. 3219-3228.
Strle K., Zhou J.H., Shen W.H. et al. Interleukin-10 in the brain // Crit. Rev. Immunol. 2001. Vol. 21. № 5. P. 427-449.
Szelenyl J. Cytokines and the central nervous system // Brain Res. Bulletin. 2001. Vol. 54. № 4. P. 329-338.
Tian L., Ma L., Kaarela T., Li Z. Neuroimmune crosstalk in the central nervous system and its significance for neurological diseases // J. Neuroinflam. 2012. Vol. 9. P. 1-10.
Tukhovskaya E.A., Turovsky E.A., Turovskaya M.V et al. Interleukin-10 increases resistance to brain ischemia through modulation of ischemia-induced intracellular Ca2+ response // Neurosci. Lett. 2014. Vol. 571. P. 55-60.
TurovskayaM.V., TurovskyE.A., Zinchenko V.P., LevinS.G., Godukhin O.V. Interleukin-10 modulates [Ca2+] i response induced by repeated NMDA receptor activation with brief hypoxia through inhibition of In- sP3-sensitive internal stores in hippocampal neurons // Neurosci. Lett. 2012. Vol. 516. № 1. P. 151-155.
TurovskyE.A., TurovskayaM.V., KononovA.V., Zinchenko VP. Short-term episodes of hypoxia induce posthypoxic hyperexcitability and selective death of GABAergic hippocampal neurons // Experim. Neurol. 2013. Vol. 250. P. 1-7.
VezzaniA., AronicaE., MazaratiA., Pitman Q.J. Epilepsy and brain inflammation // Exper. Neurology. 2013. Vol. 244. P. 11-21.
Viviani B., Bartesaghi S., Gardoni F. et al. Interleukin-1b enchances NMDA receptor-mediated intracellular calcium increases through activation of the Src family of kinases // J. Neurosci. 2003. Vol. 23. № 25. P. 8692-8700.
Zhou Z., PengX., Insolera R., Fink D.J., Mata M. Interleukin-10 provides direct trophic support to neurons // J. Neurochem. 2009. Vol. 110. № 5. P. 1617-1627.
Еще по теме Заключение:
- Заключение
- Заключение
- Заключение
- Заключение
- Заключение
- Заключение
- Заключение
- Заключение
- 2.5. Рациональное трудоустройство больных по заключению КЭК
- Патоморфологическое заключение
- Вместо заключения
- Параграф пятый. Об общих заключениях по действиям мозга
- Механизмы заключения договоров
- Заключение: от понимания к действию
- Параграф одиннадцатый. Заключения по форме головы
- Параграф четвертый. Заключения по симптомам лихорадок
- Заключение
- Заключение