12.6.1. Мускариновый холинорецептор
Среди отличительных особенностей мускаринового рецептора следует отметить не только длительность его ответа на АХ, но и быстроту возникновения этого ответа [Greengard, 1976]. Этот рецептор выделяют экстракцией из мозга быка, при этом берут только каудатные ядра, содержащие основную часть АХ мозга.
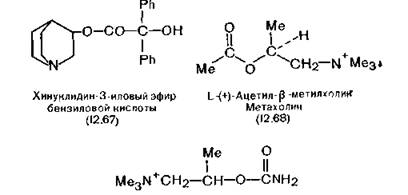
Эти ядра являются частью полосатого тела, структуры лимбической системы. Ткань этих ядер экстрагируют водным раствором холата натрия, и после центрифугирования разделяют на фракции электрофорезом. Затем добавляют [3Н]-хинук- лидин-3-иловый эфир бензиловой кислоты (12.67), специфично связывающийся с мускариновыми рецепторами. (Это соединение появилось в результате поиска специфичных для мускаринового рецептора эфиров холина.) Вначале был найден холи- новый эфир бензиловой кислоты (метацин), а соединение (12.67) можно рассматривать как его производное, так как структура холина входит в структуру хинуклидинола. После очистки комплекса реагент отмывают диализом и выделяют чистый мускариновый рецептор. Мускариновые антагонисты (атропин, гиосцин, изопропамид) блокируют связывание соединения (12.67) с рецептором, тогда как никотиновые антагонисты (тубокурарин, гексаметоний) и агонист (никотин) на связывание не влияют [Carson, 1982; Yamamura, Snyder, 1974].Мускариновый рецептор выделен также из головок фруктовой мушки Drosophila [Dudai, Ben-Barak, 1977], но в головках обычной мухи — совершенно другой тип холинорецепторов
(разд. 12.6.2).
Бетанехол (катион)
(12.69)
Большинство сведений о структуре мускаринового рецептов ра получено при изучении связи структура — активность агонистов, имитирующих действие АХ в постганглионарных синапсах. Эти рецепторы традиционно называются мускариновыми, однако следует помнить, что это название чисто историческое, так как мускарин в медицине для лечения больных не применяется.
Антагонисты мускариновых рецепторов очень сильно различаются по структуре и поэтому среди них трудно установить связь структуры с активностью. Однако из данных о действии антагониста пирензипина (бензодиазепинона) можно сделать вывод, что существуют два подтипа мускаринового рецептора.Наиболее типичный мускариновый агонист — метахолин (12.68), а-ацетил-^-метилхолин. Эта простая молекула, отличающаяся от АХ наличием дополнительной метильной группы, обладает такой же мускариновой активностью, что и природный медиатор, но совершенно не проявляет никотиноподобноё
действие. Следует отметить, что L-изомер в 200 раз активнее D-изомера [Ellenbroek, Van Rossum, 1960]. В медицинской практике метахолин в основном применяют в послеоперационном периоде для понижения тонуса кишечника. Чистым мускариновым агонистом является и бетанехол (12.69), гибрид метахолина и карбахолина (2.11), применяющийся в клинике по тем же показаниям, что и метахолин. Эти лекарственные вещества обладают важным преимуществом перед АХ: они значительно медленнее гидролизуются АХЭ.
Равновесное расстояние между четвертичным атомом азота в молекуле АХ и отрицательно заряженной группой рецептора рассчитано по разности свободных энергий взаимодействия рецептора с ацетилхолином (12.65) и с диметилбутилацетатом (12.66): оно равно 0,329 нм. Диметилбутилацетат имеет не обладающую основными свойствами головку, изостеричную катионной головке АХ. Это расстояние практически равно расстоянию максимального сближения, найденному на молекулярных моделях [Burgen, 1965].
В последние годы структура многих жестких аналогов АХ была определена рентгеноструктурным анализом для предсказания: а) конформации молекулы АХ (из множества возможных для такой гибкой молекулы), взаимодействующей с данным рецептором, и на основании этого б) стереохимии самого рецептора. Такой подход к установлению структуры рецептора достаточно дискуссионен, так как известна способность рецепторов деформироваться при взаимодействии с агонистами (разд.
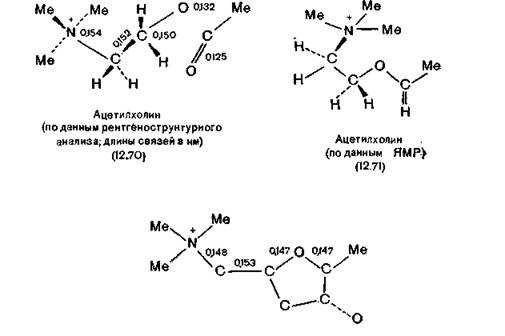
12.3).Формулой (12.70) представлена двумерная проекция молекулы ацетилхолинбромида по данным рентгеноструктурного анализа [Сапера, Pauling, Sorum, 1966]. Межатомное расстояние между N-метильной группой и эфирным атомом кислорода (т. е. атом кислорода, связанный с двумя атомами углерода) равно 0,302 нм, а между атомом азота и эфирным атомом кислорода 0,329 нм (оба эти расстояния короче, чем обычно). Оба атома кислорода находятся в одной плоскости с ближайшими тремя атомами углерода, расположенными в правой части формулы (12.70). Таким образом, геометрии молекул ацетилхолина и мускарина (12.72) [рентгеноструктурные данные Jellinek, 1957] в твердом состоянии похожи. Однако в водном растворе конформация молекулы АХ может быть совершенно иной, что обусловлено отсутствием влияния соседних молекул того же самого вещества. Поэтому конформации АХ изучали по анализу вицинальных констант в спектрах ЯМР в растворе D2O [Culvenor, Ham, 1966]. Полученные результаты подтверждают данные рентгеноструктурного анализа, за исключением того, что сложноэфирная группа находится в конформации, более свойственной сложным эфирам. Гош-конфор- мация цепи +NCCO, существующая во всех трех структурах (12.70), (12.71) и (12.72), является предпочтительной для мно
гих 1,2-дизамещенных этанов в растворе. Таким образом, предпочтительная конформация АХ весьма обычна; однако взаимодействие с рецептором может изменить ее полностью.
Мускарин
(поданнымрентгеноструктурного анализа)
(12.72)
Жесткая циклическая структура молекулы мускарина позволяет получить более полную информацию о структуре мускаринового холинорецептора, чем гибкая молекула АХ [Waser, 1961]. Из семи стереоизомеров мускарина только природный L-( + )-изомер обладает высокой ацетилхолиноподобной активностью. Атом азота должен быть четвертичным, а атом кисло-’ рода в цикле нельзя заменить даже на серу, иначе активность соединения пропадает.
На основе этих данных Waser постулировал связывание мускарина с рецептором посредством атомов азота и кислорода, входящего в цикл. Конформационные возможности молекулы мускарина значительно меньше, чем у молекулы АХ, в нем подвижна лишь боковая цепь с триметил- аммонийной головкой [Waser, 1961].Величины отношений активности L-мускарина к активности АХ для разных постганглионарных холинорецепторов изменяются от 0,1 до 5,4. Из возможных изомеров и аналогов мускарина активностью обладают лишь те, в которых метальная, гидроксильная и ониевая группы (боковой цепи) расположены так же, как и в молекуле мускарина.
Какие еще структурные модификации молекул АХ и мускарина можно проводить, сохраняя при этом сильную ацетилхолиноподобную активность? Все эти данные являются лишь по- луколичественными, и для их уточнения требуется проведение
дополнительных исследований. Дело в том, что «активность» каждого соединения складывается из двух составляющих: эффективности и сродства (разд. 7.5.2). Так, катион диметилами- ноэтилацетата (12.73), нечетвертичного аналога АХ (12.65), практически не обнаруживает мускариновой активности. В действительности же его внутренняя активность выше, чем у АХ, а низкая активность обусловлена малым сродством — в 1000 раз меньшим, чем у АХ [Gloge, Liillmann, Mutschler, 1966].
Особый интерес представляет 2-ацетоксициклопропилтри- метиламмоний-йодид (12.74), так как в этом соединении структура N—С—С—О АХ жестко закреплена ковалентными связями. Это соединение было получено в виде смеси четырех изомеров, конфигурация которых была определена рентгеноструктурным анализом [Chothia, Pauling, 1970].
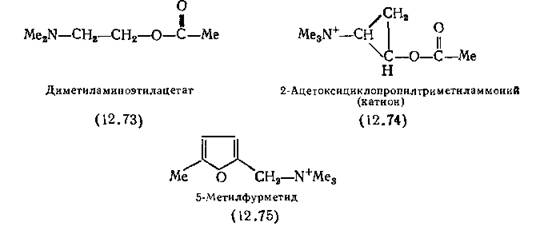
Полной мускариноподобной активностью обладает (+)- транс-изомер, остальные три малоактивны. (Активный изомер обладает лишь 1 % активности ацетилхолина, остальные изомеры еще менее активны.) Поэтому был сделан вывод, что по крайней мере мускариновая активность АХ связана с конформацией, похожей на таковую активного (IS, 2S)-изомера соединения (12.74).
5-Метил-2-триметиламмониометилфуран(12.75) (5-метилфурметид) обладает такой же мускариновой активностью, как и АХ, и почти не обладает никотиновой активностью [Armitage, Ing, 1954]. Подобно мускарину это соединение имеет в молекуле жесткий плоский цикл, но (благодаря двойным связям)—СНг-группа фрагмента —СН2—+ЫМез жестко закреплена в плоскости цикла, и поэтому форма молекулы четко определена. Следует отметить, что в отличие от мускарина в соединении (12.75) отсутствует гидроксильная группа и стереоизомерия здесь невозможна. Из данных кристаллографических исследований молекулы 5-метилфурметида [Baker et al., 1971] видно, что, хотя строение некоторых фрагментов этой молекулы совпадает со строением молекул мускарина и АХ, в другом она сильно от них отличается, поэтому эти
данные ничего не дали для выяснения структуры мускарин" вых холинорецепторов. Удаление метальной группы из соединения (12.75) резко снижает мускариновую активность, так как приводит .к понижению электронной плотности на эфирном атоме кислорода, но не меняет конформацию. Из этого можно сделать вывод, что для проявления мускариновой активности значительно важнее распределение электронов в молекуле, чем ее конформация. Как фурметид, так и 5-метилфурметид служат примерами, подтверждающими правило «пяти атомов боковой цепи» (см. ниже).
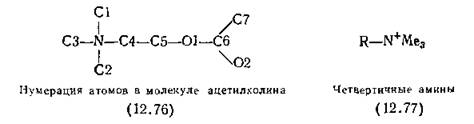
Синтез жестких агонистов АХ — задача довольно сложная. Обширные конформационные возможности молекулы АХ (12.76) зависят от четырех торсионных углов: С5—С4—N—СЗ, 01—С5—С4—N, С6—01—С5—С4 и 02—С6—01—С5. По аналогии со всеми похожими структурами первый из этих углов, вероятнее всего, постоянен — это анти-планарная вытянутая цепочка с торсионным углом 180°. Причиной этой жесткости являются стерические препятствия, создаваемые N-метильными группами, так как их ван-дер-ваальсовы радиусы мешают более плотной упаковке (разд. 8.0). Из-за планарности эфирных групп (связь С6—01) фиксирован и последний из этих углов (02—С6—01—С5). Рентгеноструктурные исследования кристаллов различных агонистов АХ показали, что два других торсионных угла могут изменяться в широких интервалах. Одна из групп исследователей пришла к выводу, что для проявления мускариновой активности необходимо только расположение атомов С1 и С7 АХ по одну сторону молекулы [Baker et at, 1971]. Другая группа на основании похожих доказательств (в сочетании с данными для антагонистов, менее значимыми) постулировала, что АХ взаимодействует с мускариновыми рецепторами эфирным атомом кислорода и аммонийным катионом [Beers, Reich, 1970].
Еще по теме 12.6.1. Мускариновый холинорецептор:
- 12.6. Холинорецепторы
- Никотиновые холинорецепторы
- Третий тип холинорецепторов.
- Д. Антагонисты мускариновых рецепторов.
- Профиль безопасности ТЦА
- Механизм токсического действия
- Экстракардиаьные причины.
- 7.4. Патогенез аллергического ринита.
- 1.4. Антидоты Би-Зет
- Механизм токсического действия
- По токсичности ФОС разделяют на 4 группы.
- Отравляющие и высокотоксичные вещества судорожного действия Конвульсанты, действующие на холинореактивные синапсы
- Глава IV. Характеристика спектра карДиоспецифических аутоантител у Детей с нарушением ритма серДца и провоДимости и без аритмии.
- Состояние основных нейротрансмиттерных систем при развитии БА
- Глава 25. Миастения
- Б. Изменение катионной головки ацетилхолина.
- Морфофункциональные изменения в поле CA1 гиппокампа крыс при использовании пентилентетразоловой и литий-пилокарпиновой моделей судорожных состояний
- Г. Оптимальное положение атома кислорода в боковой цепи.
- А н т и х о л и н е р г и ч е с к и е п р е п а р а ты.
- В. Правило «пяти атомов в боковой цепи».