Никотиновые холинорецепторы
Никотиновый холинорецептор в отличие от мускаринового значительно быстрее реагирует на появление стимула. Он был выделен из электрических органов рыб (разд. 2.1, рис. 2.1, том 1).
Этот гликопротеин можно пометить змеиным токсином и а- бунгаротоксином, обычно используемым в виде [3Н]-ацетил- производного. Реагент специфично связывается с ацетилхолин-связыйающим местом рецептора. Такой комплекс токсин — рецептор можно использовать для очистки рецептора, отделяя токсин в, конце очистки длительным диализом. В 1980 г. очищенный рецептор был встроен в искусственную плоскую липидную мембрану с целью демонстрации его пермеазной природы. С помощью дифракции рентгеновских лучей было установлено,, что рецептор пронизывает мембрану, выступая понемногу с каждой стороны. Диаметр рецептора равен приблизительно» 8,5 нм, и он включает в себя ионный канал диаметром 0,65 нм. Такой реконструированный рецептор под действием АХ увеличивал проницаемость мембраны для катионов, причем эффект- АХ блокировался тубокурарином [Nelson et al., 1980].
Каждая рецепторная розетка (см. рис. 2.1) включает в себя пять параллельных цепочек, образующих ионный канал. На двух цепочках, называемых а-субъединицами, расположены места связывания ацетилхолина. Остальные три играют чисто структурную роль[7] [8]. На выделенной из Torpedo californica иРНК, кодирующей а-субъединицу, была синтезирована и проклонирована комплементарная ДНК- Так была получена иРНК в количествах, достаточных для установления нуклеотидной последовательности, по которой в свою очередь была определена первичная структура а-субъединицы с ОММ 50116, содержащей 437 аминокислотных остатков. Дисульфидный мостик около ацетилхолинсвязывающег» места, по-видимому, образован двумя остатками Cys-128 и 142. Что касается самого ацетилхолинсвязывающего места, то предполагается, что в его состав входят остатки Asp-138 (или Clu-129) и His-134. Для возникновения физиологического ответа как минимум две молекулы АХ должны связаться с рецептором. Выброс АХ, длящийся 1 мкс, вызывает в скелетной мышце ответ продолжительностью не менее 10 мкс. Судя по этим цифрам, открывание ионного канала контролируется непосредственно рецептором и для этого не требуется предварительного фосфорилирования. Таким образом, механизм никотинового ответа является более простым и более прямым, чем постулированный Greenyard механизм мускаринового ответа (разд. 12.6.1). Все приведенные выше данные получены для рецептора, выделенного из электрического органа рыб, однако сходный белок был выделен из скелетных мышц млекопитающих [Dolly, Barnard, 1977; Froehner, Reiness, Hall, 1977] и из коры головного мозга морских свинок. Последний имеет ОММ около» 86 000, а связывание с ним [3Н]-бунгаротоксина игибируетсяі тубокурарином и галламином [Bosnian, 1972]. Более подробно о никотиновых рецепторах см. Karlin /(1980) и Conty-Tronconi, Raferty (1982). Задолго до выделения никотинового рецептора было начато ■изучение связи структура — активность в ряду агонистов, действующих на активируемые АХ места, традиционно называемые никотиновыми. Этот термин, так же как и термин «мускариновый», имеет чисто историческое значение, так как никотин клинического применения не имеет. Известно, что никотин проявляет холиномиметические свойства у позвоночных в трех структурах: а) на моторной концевой пластинке произвольной мышцы в нервно-мышечном соединении; б) в ганглиях симпатических и парасимпатических нервов и в) в коре головного мозга. Наиболее доступна для действия никотина структура t(a) (это следует хотя бы из того, что никотин является ядом для млекопитающих). Однако у насекомых, в нервно-мышечных соединениях которых не содержится АХ, основное действие никотина сосредоточено в ганглиях ЦНС [Yaeger, Munson, 1945]. В настоящее время никотин мало используется в качестве инсектицида. Простейшее соединение, обладающее сильным никотиновым ■(но не мускариновым) действием, основной агонист — это тет- раметиламмоний-хлорид (12.77, R=Me). В ряду Н-холиноми- метиков это такое же стандартное соединение, как и метахо- дин (12.68) в ряду М-холиномиметиков. По силе действия на автономные ганглии соли тетраметил- аммония не отличаются от АХ [Burns, Dale, 1915], однако на нервно-мышечное соединение скелетных мышц (никотиновая активность) они действуют в сто раз слабее, чем АХ, а на ■соединение с гладкими мышцами (мускариновая активность) — в тысячу раз слабее. Взаимодействие медиатора с автономными ганглиями кратковременно, а в нервно-мышечном соединении этот процесс более длителен, и для него предпочтительнее молекулы, способные образовывать водородные связи, например, как АХ — за счет кето-атома кислорода. Очевидно, катион тетраметиламмония определяет биологическую активность АХ; при необходимости длительного взаимодействия важную роль играет и остальная часть молекулы, обеспечивая дополнительное связывание с рецептором. Из соединений ряда катионов алкилтриметиламмония (родоначальником которого является тетраметиламмоний) максимальной активностью обладают соли н-пентилтриметиламмо- ния, их действие на никотиновые рецепторы такое же, как у никотина, и в восемь раз сильнее, чем у АХ [Willey, 1955]. Молекулы никотиновых агонистов, вероятно, должны находиться в гош-конформации (определение см. разд. 12.3). Установлено, что в кристаллах и растворах большинство этих агонистов действительно имеют гош-конформацию (синклинальную) фрагмента —+N—С—С—О [Baker et al., 1971; Calvenor, Ham, 1966; Сапера et al., 1966]. Исключение составляет карбахолин (2.11), существующий в кристаллах в вытянутой (антипланар- ной) конформации, а в растворе — в гош-конформации [Ваг- rans, Clastre, 1970; Baker et al., 1971]. Однако данные по конформации агонистов, содержащих фрагменты —+N—С—С—S— и —+N—С—С—Se—, для которых вытянутая (антипланарная) конформация является преимущественной как в кристаллах [Shefter, Mautner, 1969], так и (по данным ЯМР) в растворах [Cashley, Mautner, 1970], ставят под сомнение правомерность гош-гипотезы. Например, тиохолин и S-метилтиохолин (оба в вытянутой конформации) — сильные агонисты, тогда как их кислородные аналоги холин и О-метилхолин (оба в гош-конформации) не активны [Mautner, Bartels, Webb, 1966]. Более того, замена карбонильного атома кислорода в молекуле АХ атомом серы не приводит к изменению гош-конформации [Mautner, Dexter, Low, 1972] и как сам АХ, так и ацетилтиохолин (12.87) высокоактивны в различных биологических тестах [Scott, Mautner, 1967; Mautner et al., 1966; Mautner, 1969]. Эти данные свидетельствуют о том, что, вероятно, для проявления агонистической активности исходная конформация не менее важна, чем распределение электронов. При изучении рецепторного связывающего места был применен иммунологический подход, для чего были получены кроличьи антитела к фениловому эфиру холина (12.86). К сожалению, при попытке использования этих антител в качестве модели никотинового рецептора выяснилось, что они не способны различать ни вещества мускаринового и никотинового типа действия, ни агонисты и антагонисты [Marlow, Metcalf, Burgen, 1969]. Антагонисты никотиновых рецепторов. Ни один чистый никотиновый агонист не используется в медицине. Карбахолин (2.11) обладает не только никотиновыми, но и выраженными мускариновыми свойствами. Антихолинэстеразные лекарственные вещества усиливают действие эндогенного АХ, что означает одновременную активацию мускариновых и никотиновых структур. В качестве примера можно привести использование физостигмина (2.8) для лечения острой глаукомы, фосфорорганических соединений (разд. 13.3)—для лечения хронических случаев этого заболевания, часто встречающегося у пожилых людей, и прозерина (2.10) —для снятия нервно-мышечного блока при миастении гравис. Последний лекарственный препарат обладает слабым никотиновым действием и на произвольные мышцы. Хотя стимуляцию никотиновых рецепторов мало используют в терапии, в хирургии широко применяют никотиновые антагонисты, позволяющие добиваться глубокого мышечного расслабления и снижения доз общих анестетиков. В 1942 г. для этой цели был применен тубокурарин (2.6), затем в дополнение к нему появились другие четвертичные аммониевые катионы. В настоящее время известны два основных типа таких лекарственных веществ. Представители первого типа, например тубокурарин, препятствуют деполяризации мембран ацетилхолином, но сами не оказывают на них никакого ацетилхолиноподобного действия. Они являются, конкурентами АХ при его взаимодействии с рецептором, но не препятствуют его синтезу, высвобождению и распаду. Таблица 12.3. Зависимость специфичности действия от длины цепи бис- триметиламмоиийных катионов на два типа холинорецепторов (антиникотиновое действие) Нерв — мышца 0,07 Примечание. Приведены относительные активности, для каждого типа макси- мальная величина активности 1,0 [Paton, Zaimis, см. текст]. ладающие достаточно высокой ацетилхолиноподобной активностью для того, чтобы вызвать деполяризацию до того, как наступит нервно-мышечный блок. Триметиламмонийные группы этих соединений не испытывают стерических затруднений, поэтому возможно взаимодействие по меньшей мере одного конца молекулы с анионным местом холинорецептора, хотя вся молекула в целом достаточно велика, чтобы закрыть полость. Сродство этих бис-ониевых солей к трем основным типам холинорецепторов отличается высокой специфичностью. Все они практически не действуют на постганглионарные рецепторы, однако их активность в ганглиях и нервно-мышечных соединениях может быть исключительно высокой при условии, что для каждого из этих двух видов рецепторов выбран надлежащий гомолог. Декаметоний в дозах, в сто раз превышающих дозы, полностью блокирующие нервно-мышечное соединение, совершенно не действует на ганглии. Гексаметоний, наоборот, в стократной дозе по сравнению с дозой, блокирующей ганглии, не действует на нервно-мышечное соединение [Paton, Zaimis, 1949, 1952]. Эта специфичность проиллюстрирована данными в табл. 12.3. Ганглиоблокаторы типа гексаметония (7.30) вошли в медицинскую практику около 1950 г. в качестве препаратов для лечения гипертонии. Однако их действие на организм оказалось настолько неспецифичным, с множеством побочных эффектов, что в последствии эти препараты были вытеснены ^-адрено- блокаторами и лекарственными веществами, расслабляющими мышцы непосредственно в малых кровеносных сосудах. Лекарственное вещество, способное связываться одновременно с двумя местами связывания на рецепторе, должно отличаться очень высоким сродством к последнему, так как: а) если один конец молекулы в результате диссоциации удаляется от поверхности, то другой конец будет ее удерживать на близком расстоянии от поверхности и тем самым способствовать рекомбинации и б) молекулы лекарственного вещества будут соединены более прочно за счет ван-дер-ваальсовых сил с частью биологической поверхности, лежащей между двумя местами связывания. Введение эфирной группы в молекулы бис-ониевых блокаторов нервно-мышечной передачи резко повышает их способность ингибировать деполяризацию, как, например, холино- вые эфиры дикарбоновых кислот типа (12.88). Из этих соединений в хирургии наиболее широко применяют дитилин (7.29) из-за его способности медленно гидролизоваться холинэстеразой плазмы, что способствует прекращению действия препарата на больного после операции. (В разд. 3.5.4 упоминалось новое лекарственное вещество атракурий, гидролизующийся под действием воды.) Усиление действия соединений типа (12.88) при введении эфирной группы указывает на наличие в рецепторе групп, способных к образованию водородных связей. В ряду подобных соединений вторая триметиламмонийная группа может быть без ущерба для действия заменена атомом водорода [Danilov et al., 1974]. Замена одной триметиламмонийной группы в молекуле декаметония на другую гидрофильную (например, метокси) группу не нарушает действие вещества и, следовательно, для связывания никотиновых антагонистов необходима лишь одна кислотная группа в рецепторе. О расстоянии между двумя местами связывания в холино- рецепторе можно судить по размерам молекулы панкуроний- бромида (12.89), сильного неполяризующего миорелаксанта, применяющегося в клинической практике [Baird, Reid, 1967]. Жесткость его молекулы была подтверждена спектрами ЯМР в растворе. По данным рентгеноструктурного анализа расстоя- 248 ние между двумя атомами азота в молекуле панкурония равно 1,108 нм, что сравнимо с таковым у Ь^О.О'-триметил-сІ-тубоку- рарин-дийодида в кристаллах, равным 1,07 нм (вероятно, таково же расстояние между атомами азота и в самом тубокура- рине). Следует отметить, что панкуроний структурно ближе АХ, чем соединения бис-ониевого тина, так как в его молекуле дважды повторяется фрагмент Me—С (О)—О—CHR—CHR— —N+R3 [Savage et al., 1970]. Молекула декаметония (7.28) в кристаллическом состоянии полностью вытянута и атомы азота удалены друг от друга на расстояние 1,37 нм [Lonsdale, Milledge, Pant, 1965], однако в растворе это расстояние, видимо, меньше. А у молекулы дитилина, гибкой, как и у всех деполяризующих агентов, расстояние между атомами азота в кристаллическом состоянии зависит от аниона, образующего данную соль, и варьируется от 0,78 до 1,19 нм. Бис-ониевые соединения, содержащие между атомами азота цепочку из 16 атомов (не обязательно углерода), обозначают N-16-N, или «С-16». Примеры таких соединений — суберилди- холин (субехолин), себацинилдихолин и карболоний-бромид (12.90), изредка применяемый в хирургии. Холинорецептор, чувствительный к соединениям «С-16», появился на ранней стадии эволюции и широко распространен в животном царстве. Нервно-мышечный синапс моллюсков, иглокожих и первичнохордовых ингибируется соединениями «С-16», но не «С-10»; некоторые аннелиды чувствительны к соединениям «С-10», однако все черви — к соединениям «С-16». Амфибии значительно более чувствительны к «С-16»-, чем к «С-10»-ингибиторам, а высшие позвоночные примерно одинаково чувствительны к обоим типам. Из того факта, что невозможно блокировать рецепторы для соединений структуры «С-16», не заблокировав рецепторы для соединений структуры «С-10» (и наоборот), следует, что эти рецепторы должны иметь общие группы [Khromov-Borisov, Michelson, 1966; Михельсон, Земаль, 1970]. На основании этих данных была создана тетрамерная модель мышечного холинорецептора, согласно которой четыре анионных связывающих места расположены в вершинах квадрата. Соединения «С-10» взаимодействуют с двумя центрами, расположенными на одной стороне квадрата (например, дитилин), а соединения «С-16» — с центрами на диагонали [Khromov- Borisov, Michelson, 1966] *. Об истории создания и применении никотиновых антагонистов см. разд. 7.3. 1 Более подробно об исследовании структуры холинорецептора химикофармакологическими методами см. Н. В. Хромов-Борисов в ки.: Биохимическая фармакологии (под ред. П. В. Сергеева). М., Медицина, 1982. — Примеч. ред.
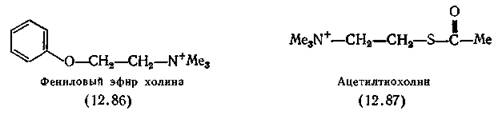
Тип холинергического синапса Число метиленовых групп 4 5 6 7 8 9 10 12 Нерв — нерв 0,01 0,8 1,0 0,1 0,02 0,7 0,1 0,55 0,2 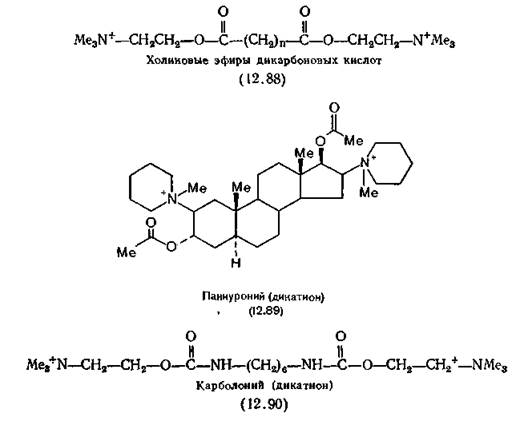
Еще по теме Никотиновые холинорецепторы:
- 12.6. Холинорецепторы
- Третий тип холинорецепторов.
- Никотиновые ацетилхолиновые рецепторы: физиологические функции и роль в нейродегенеративных процессах
- 12.6.1. Мускариновый холинорецептор
- Общие данные о никотиновых ацетилхолиновых рецепторах: структура, подтипы, локализация
- Механизм токсического действия
- Нарушения обмена триптофана.
- Г. Оптимальное положение атома кислорода в боковой цепи.
- Экстракардиаьные причины.
- 1.4. Антидоты Би-Зет
- Механизм токсического действия
- По токсичности ФОС разделяют на 4 группы.
- Отравляющие и высокотоксичные вещества судорожного действия Конвульсанты, действующие на холинореактивные синапсы