Распространение метилирования ДНК.
Метилирование оснований в ДНК было открыто свыше 50 лет назад и наблюдается практически во всех классах живых организмов. ДНК прокариот содержит модифицированные основания ^-метиладенин и 5-метилцитозин, тогда как для эукариот характерно наличие в основном 5-метилцитозина.
Он присутствует в ДНК грибов и растений (Finnegan et al. 2000; Martienssen and Colot 2001). В царстве животных наблюдается широкий спектр уровня метилирования генома. На одном из полюсов располагается нематода Caenorhabditis elegans, чей геном лишен 5-метилцитозина и не кодирует соответственно ДНК-метилтрансферазы. Рядом с ней располагается плодовая мушка Drosophila melanogaster. Долгое время считали, что ее геном также не подвергается метилированию. Однако у нее имеется ген, кодирующий белок, гомологичный ДНК-метилтрансферазам млекопитающих (Hung et al. 1999; Tweedie et al. 1999). Недавно было показано, что 5-метилцитозин в ДНК D. melanogaster присутствует в очень малых количествах (Gowher et al. 2000; Lyko et al. 2000). На противоположной стороне от C. elegans находятся позвоночные животные. Их геном имеет самый высокий уровень 5-метилцитозина во всем животном царстве. Метилирование ДНК у позвоночных настолько широко распространено в геноме, что говорят о его тотальном метилировании.У млекопитающих 5-метилцитозин в геноме встречается практически только в составе CpG динуклеотида. При этом количество CpG динуклеотидов снижено из-за высокой мутабильности 5-метилцитозина, который с высокой вероятностью подвергается спонтанному дезаминированию и превращению в тимин. Это обстоятельство привело в процессе эволюции к многочисленным заменам пар G:C на A:T, в результате чего в геноме млекопитающих динуклеотидов CpG в ~ 5-раз меньше, чем следовало бы ожидать (Gardiner-Gardner and Frommer 1987). У человека
~80% всех CpG динуклеотидов метилировано, и эти метилированные CpG динуклеотиды рассредоточены по геному в виде одиночных сайтов (Bird
1995).
В то же время в геноме позвоночных существуют скопления неметилированных CpG динуклеотидов, именуемых CpG-островками (у человека ~20% от всего числа CpG динуклеотидов). Эти последовательности обладают длиной от 0.5 до 5 т.п.н. (в среднем 1 т.п.н.), встречаются приблизительно один раз на 100 т.п.н., имеют высокую плотность CpG динуклеотидов (близкое к расчетному 1:16), их GC состав превышает 0.55, а отношение экспериментально найденного числа CpG к теоретически возможному составляет не менее 0.6 (Antequera and Bird 1993; Takai and Jones 2002). На основании компьютерного анализа в геноме человека предсказано существование 29 тыс. CpG-островков (Lander et al. 2001; Venter et al. 2001). CpG-островки расположены вблизи генов, преимущественно в 5' области (регуляторные последовательности, промоторы, последовательности первого экзона). Считается, что ~60% человеческих генов содержат CpG-островки (Antequera and Bird 1993). Они имеются, по-видимому, у всех генов, выполняющих базовые клеточные функции (гены "домашнего хозяйства"), и у ~40% генов, выполняющих специализированные функции (тканеспецифические гены). Большинство из CpG-островков остаются неметилированными на всех стадиях развития организма и во всех типах тканей, даже когда ассоциированный с ними ген не функционирует. Например, тканеспецифические гены человека а-глобин (Bird et al. 1987) и а2(1)-коллаген (McKeon et al. 1982) имеют CpG-островки, которые остаются неметилированными во всех протестированных тканях, независимо от статуса экспрессии гена. В то же время в клетке может происходить метилирование CpG-островка, что сопровождается стабильной инактивацией связанного с ним гена. Так, во время развития метилирование CpG-островков наблюдается при установлении геномного импринтинга и инактивации Х хромосомы (см. обзор Bird 2002), а также наблюдается при патологических
процессах, например при канцерогенезе.
Считается, что метилирование CpG-островков в регуляторных областях
приводит к перестройке хроматина в неактивное состояние, что вызывает
инактивацию транскрипции метилированного гена (см.
обзор Baylin and Herman 2000; Jaenisch and Bird 2003). Показано, что важную роль в этом играет семейство метил-CpG-связывающих белков, привлекая кметилированному локусу ДНК гистоновые деацетилазы, которые, в свою очередь, удаляют ацетильные группы у гистонов в составе нуклеосом. Такая
модификация гистоновых белков приводит к превращению открытой, транскрипционно-активной структуры хроматина в закрытую,
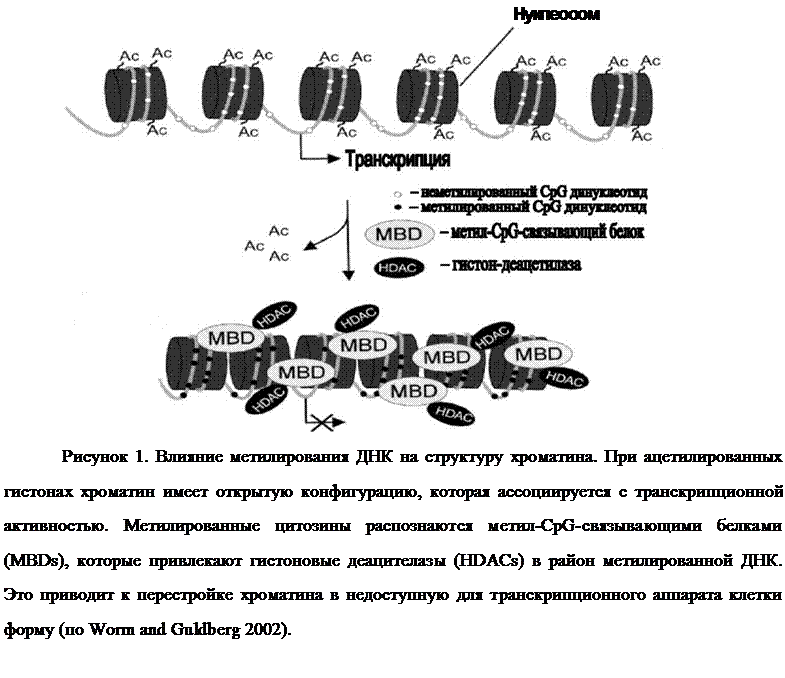
транскрипционно-неактивную (рис. 1).
В тоже время транскрипция может регулироваться метилированием одиночных CpG динуклеотидов, расположенных в области промотора в зоне сайта связывания ряда транскрипционных факторов. Метилирование в этом случае имеет относительно локальный характер, препятствуя связыванию чувствительных к метилированию факторов транскрипции, и варьирует в широких пределах в разных тканях. В настоящий момент известно около 20 метилчувствительных транскрипционных факторов. Например, такое сайт-специфическое метилирование и ассоциированная с ним инактивация транскрипции наблюдается в случае с промотором GK-интерферона (Melvin et al. 1995) и участком связывания CREB (cyclic AMP response element-binding protein) промотора гена в-глобина (Iguchi-Ariga et al. 1989). Подобная регуляция может наблюдаться как для генов, обладающих CpG-островками, так и для тканеспецифических генов, их не имеющих. Однако роль метилирования в этом случае ограничивается влиянием на доступность сайта связывания чувствительному к метилированию транскрипционному фактору и реализуется только при наличии в клетке соответствующего белкового фактора.
Еще по теме Распространение метилирования ДНК.:
- 1.7.3. Болезнь Альцгеймера
- ОГЛАВЛЕНИЕ
- - 6 -Введение
- Распространение метилирования ДНК.
- Функция метилирования ДНК.
- Метилирование во время развития
- Ферменты метилирования
- Метилирование как динамический процесс
- Сравнительный анализ современных методов определения статуса метилирования ДНК.
- Методы идентификации CpG-островков, аберрантно-метилированных в опухолях.
- СЛОВАРЬ ТЕРМИНОВ
- КАНЦЕРОГЕНЕЗ КАК КОМПЛЕКСНОЕ ИЗМЕНЕНИЕ ГЕНОТИПА КЛЕТКИ
- Общее учение об опухолях
- ВВЕДЕНИЕ
- Молекулярно-генетические маркеры.
- Глава 4: Исследование молекулярно-генетических маркеров в неизмененной слизистой культи желудка у больных, оперированных по поводу рака в объеме резекции.
- СПИСОК ЛИТЕРАТУРЫ
- Современные представления о канцерогенезе и опухолевой прогрессии
- УКАЗАТЕЛЬ ЛИТЕРАТУРЫ
- Морфологические исследования в детской онкологии