Метилирование как динамический процесс
Статус метилирования ДНК в любой клетке устанавливается в результате сочетания динамических процессов метилирования и деметилирования. В популяции клеток CpG динуклеотиды могут иметь один из трех профилей метилирования (Turker 1999): полностью метилированные (уровень метилирования почти 100%), неметилированные (около 0%) и частично метилированные (между 0 и 100%), - представляя собой усредненный показатель метилирования, определенный анализом большого числа аллелей.
В случае импринтированных генов и генов инактивированной в женских клетках хромосомы Х промежуточный уровень метилирования (около 50%) отражает альтернативное состояние метилирования двух аллелей. Такая наследуемая совокупность всех неметилированных, частично метилированных и полностью метилированных CpG динуклеотидов для данной области ДНК получила название паттерна метилирования. В первых экспериментах, показавших существование воспроизведения статуса метилирования искусственно метилированной ДНК при делении клеток, также наблюдалось относительно низкая точность этого процесса (Wigler et al. 1981). После многих клеточных генераций, исследуемая ДНК сохранила метилирование, но на значительно меньшем уровне, чем это было в начале. Количественные исследования эндогенных CpG динуклеотидов подтвердили относительную стабильность паттерна метилирования (Riggs et al. 1998). Клеточные клоны, в которых изучаемые сайты первоначально были неметелированными, приобретали метилирование, а клоны с метилированными CpG динуклеотидами его теряли. Динамические изменения в деталях статуса метилирования исследовались в области промотора мышиного гена Aprt (Turker 1999). Этот ген обладает CpG-островком, который включает промотор, первый и второй экзоны и первый интрон. Было показано, что статус метилирования CpG динуклеотидов, расположенных в непосредственной близости от начала CpG-островка вне его границ, не однороден в клеточной популяции, тогда как CpG динуклеотиды в составе всех сайтов метилчувствительной рестриктазы HpaII внутри CpG-островка не метилированы. На основании многочисленных экспериментальных данных была предложена модель, объясняющая образование и поддержание паттерна метилирования 5' области гена Aprt (рис. 3). В раннем эмбриогенезе для 5' области гена Aprt характерно неметилированное состояние. Предполагают, что процесс метилирования19
начинается после имплантации бластоцисты de novo метилированием в так называемом центре метилирования, активную часть которого составляют две копии B1 элементов, повторяющихся последовательностей грызунов, аналогичных повторам Alu у приматов. Затем метилирование распространяется «ниже» по направлению к промотору гена.
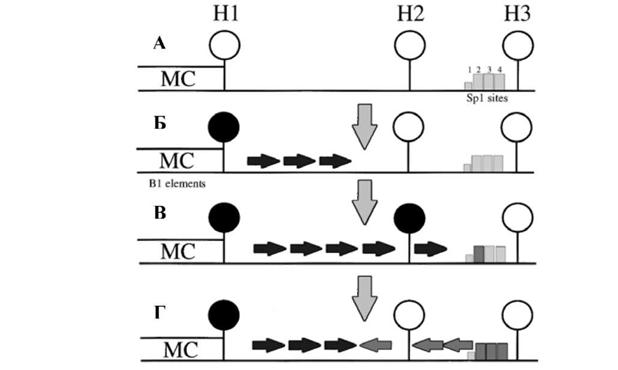
Рисунок 3. Модель возникновения и поддержания паттерна метилирования 5' области гена Aprt мыши. А. На ранних стадиях эмбриогенеза CpG динуклеотиды в составе сайтов рестрикции фермента HpaII (H1-H3) не метилированы. Б. После имплантации происходит de novo метилирование центра метилирования (МС), содержащего В1 повторы и сайт Н1, и распространение метилирования за пределы этой области. В. В некоторых случаях метилирование может распространяться вплоть до сайта Н2, расположенного в непосредственной близости от начала CpG-островка. Г. Наличие сайтов связывания транскрипционного фактора Sp1 препятствует распространению метилирования на область промотора. Такое блокирование метилирования может распространяться от области промотора в сторону центра метилирования и приводить к деметилированию сайта Н2, препятствуя поддерживающему метилированию (по Turker
1999).
Однако, сайты связывания транскрипционного фактора Sp1 предохраняют промотор от экспансии метилирования, блокируя процесс вблизи сайта метилчувствительной рестриктазы ^aII. Как показано, делеции или мутации в сайтах Sp1 приводят к метилированию CpG-островка гена (Macleod et al. 1994; Mummaneni et al. 1995). В силу конкуренции между
импульсами метилирования, исходящими из центра метилирования, и импульсами деметилирования, исходящими из структур промотора, этот сайт находится в динамически изменяющемся состоянии (метилирован менее чем на 50%). Таким образом, в установлении статуса метилирования участвуют два взаимоисключающих друг друга процесса: деметилирование ранее метилированных CpG динуклеотидов и de novo метилирование неметилированных CpG динуклеотидов. И, хотя, по всей видимости, паттерн метилирования ДНК в клетке в процессе развития передается по наследству точно (см. обзор Bird 2002), на уровне одиночных CpG динуклеотидов статус метилирования может варьировать. Поэтому представляется возможным нарушение профиля метилирования при сдвиге равновесия, вызванного усилением или ослаблением одного из двух противоборствующих процессов и развивающегося на протяжении нескольких клеточных поколений. Именно это, по всей видимости, и происходит при старении и злокачественной трансформации клетки.
Еще по теме Метилирование как динамический процесс:
- Глава 7. Бронхиальная астма
- ОГЛАВЛЕНИЕ
- Метилирование как динамический процесс
- Прионоподобные домены и их роль в обратимой и необратимой агрегации белков
- Практические рекомендации: