1.1. оПХМЖХО ЛЕРНДЮ ЛЕРХКВСБЯРБХРЕКЭМНИ ожп ЯН ЯРЮРХЯРХВЕЯЙХЛХ GC- АНЦЮРШЛХ ОПЮИЛЕПЮЛХ.
гЮДЮВЕИ МЮЬЕЦН ХЯЯКЕДНБЮМХЪ АШКН НАМЮПСФЕМХЕ CpG-НЯРПНБЙНБ, ЛЕРХКХПНБЮММШУ Б НОСУНКЪУ ЬЕИЙХ ЛЮРЙХ. дКЪ ЕЕ ПЕЬЕМХЪ ЛШ ХЯОНКЭГНБЮКХ ЛЕРНД ЛЕРХКВСБЯРБХРЕКЭМНИ ожп ЯН ЯРЮРХЯРХВЕЯЙХЛХ GC-АНЦЮРШЛХ ОПЮИЛЕПЮЛХ (яо-ожп) (Gonsalgo et al.
1997). оПХМЖХО ЛЕРНДЮ ГЮЙКЧВЮЕРЯЪ Б ЯКЕДСЧЫЕЛ. дмй ХГ ДБСУ ЯПЮБМХБЮЕЛШУ НАПЮГЖНБ (Б МЮЬЕЛ ЯКСВЮЕ ХГ НОСУНКЕБНИ РЙЮМХ Х КЕИЙНЖХРНБ ХКХ ОПХКЕЦЮЧЫЕИ Й НОСУНКХ ЛНПТНКНЦХВЕЯЙХ МНПЛЮКЭМНИ РЙЮМХ, БГЪРШУ НР НДМНЦН Х РНЦН ФЕ ОЮЖХЕМРЮ) НАПЮАЮРШБЮКХ ЛЕКЙНЫЕОЪЫЕИ ПЕЯРПХЙЖХНММНИ ЩМДНМСЙКЕЮГНИ (ПХЯ. 5ю), Б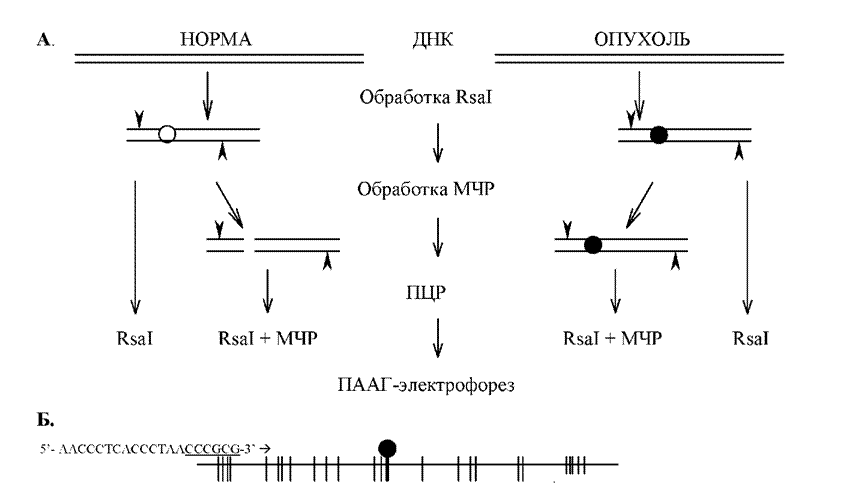
-ОПНДСЙР ожп ОЕПБНЦН РХОЮ; - ОПНДСЙР ожп БРНПНЦН РХОЮ. вХЯКЮЛХ ЯОПЮБЮ СЙЮГЮМШ ПЮГЛЕПШ ЛЮПЙЕПМШУ ТПЮЦЛЕМРНБ.
- 63 -

ТЕПЛЕМРЮЛХ, ЯНДЕПФЮЫХЛХЯЪ Б ОНКХКХМЙЕПЕ. бЯКЕД ГЮ ЩРХЛ ОПНБНДХКХ ЯХЙБЕМЯ ЙКНМНБ.
оНЯКЕ ЯЕЙБХМХПНБЮМХЪ АШК ОПНБЕДЕМ ЮМЮКХГ ОНКСВЕММШУ ОНЯКЕДНБЮРЕКЭМНЯРЕИ. бН-ОЕПБШУ, ХЯЯКЕДСЕЛШЕ ТПЮЦЛЕМРШ КНЙЮКХГНБШБЮКХ Б ЦЕМНЛЕ ВЕКНБЕЙЮ. оПХ НРПХЖЮРЕКЭМНЛ ПЕГСКЭРЮРЕ ОНХЯЙ ОПНБНДХКХ Б ЦЕМНЛЮУ ДПСЦХУ ОНГБНМНВМШУ. дКЪ ПЕЬЕМХЪ ЩРНИ ГЮДЮВХ ЛШ ХЯОНКЭГНБЮКХ МЮАНП ОПНЦПЮЛЛ ОНХЯЙЮ ЦНЛНКНЦХИ BLAST╝ (ЯЛ. ПЮГДЕК "лЮРЕПХЮКШ Х ЛЕРНДШ", О. 14.1). бН-БРНПШУ, МЕНАУНДХЛН АШКН СЯРЮМНБХРЭ, ЪБКЪЕРЯЪ КХ НАМЮПСФЕММЮЪ МЮЛХ ОНЯКЕДНБЮРЕКЭМНЯРЭ CpG-НЯРПНБЙНЛ. дКЪ ЩРНЦН ЛШ ХЯОНКЭГНБЮКХ НАЫЕОПХМЪРШЕ ЙПХРЕПХХ CpG-НЯРПНБЙНБ (ЯЛ. ПЮГДЕК "лЮРЕПХЮКШ Х ЛЕРНДШ", О. 14.2).
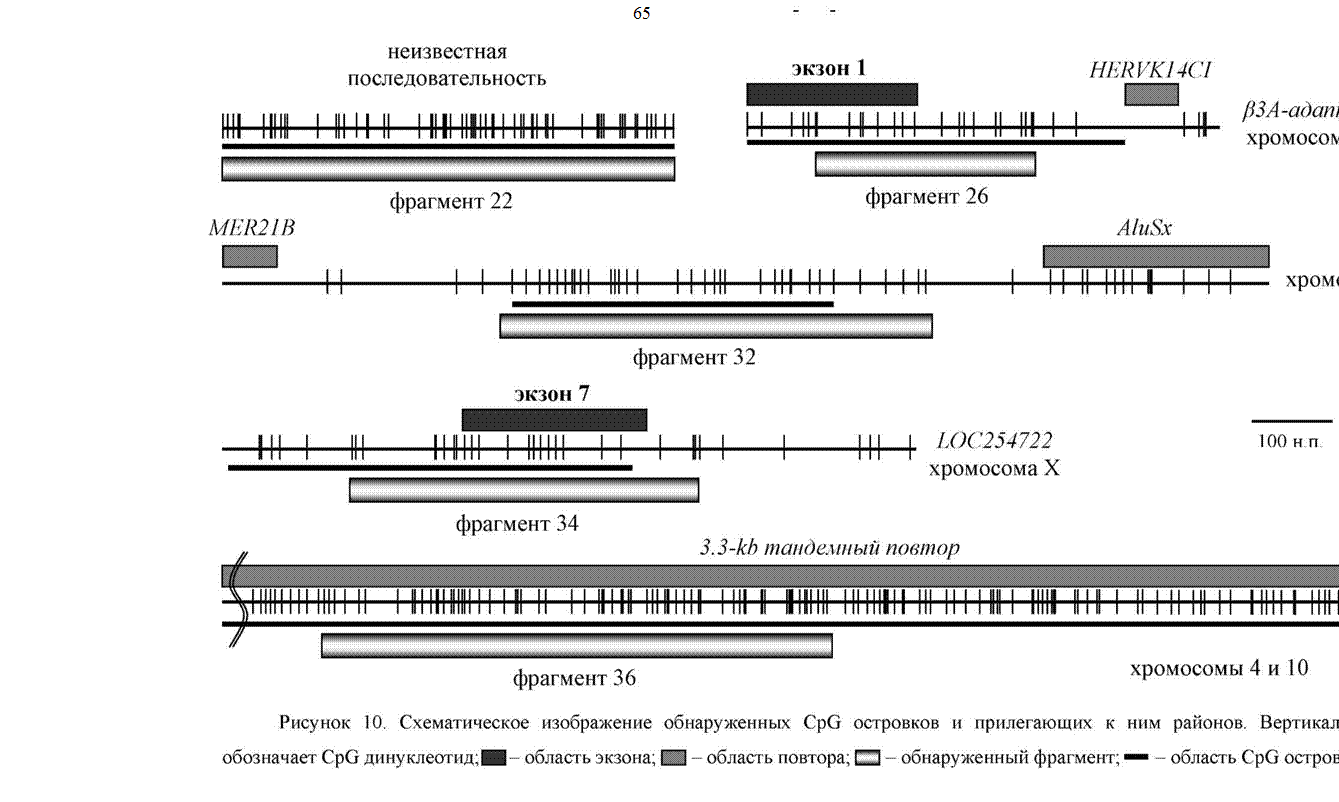
пЕГСКЭРЮРШ ЮМЮКХГЮ БШЪБКЕММШУ ТПЮЦЛЕМРНБ ОПЕДЯРЮБКЕМШ МЮ ПХЯСМЙЕ 10 Х Б РЮАКХЖЕ 4.
тПЮЦЛЕМР 18 КНЙЮКХГНБЮМ БН БРНПНЛ ХМРПНМЕ ЦЕМЮ LOC148870, ПЮЯОНКНФЕММНЦН МЮ ОЕПБНИ УПНЛНЯНЛЕ Б ГНМЕ 1p36.32 (Homo sapiens chromosome 1 reference genomic contig NT 004321, www.ncbi.nlm.nih.gov/ entrez/viewer.fcgi?val=NT_004321.12). дЮММШИ ЦЕМ ОНКСВЕМ ЮБРНЛЮРХВЕЯЙХЛ ЙНЛОЭЧРЕПМШЛ ЮМЮКХГНЛ Я ХЯОНКЭГНБЮМХЕЛ ЛЕРНДЮ ОПЕДЯЙЮГЮМХЪ BLAST.
оНДРБЕПФДЕМХЕЛ МЮКХВХЪ ЦЕМЮ ЯКСФХР ЯСЫЕЯРБНБЮМХЕ ЦНЛНКНЦХХ Я ОНЯКЕДНБЮРЕКЭМНЯРЭЧ НДМНЦН ЙКНМЮ EST. цЕМ ХЛЕЕР ОПНДСЙР -ЦХОНРЕРХВЕЯЙХИ АЕКНЙ FLJ32825, ТСМЙЖХЪ ЙНРНПНЦН МЕХГБЕЯРМЮ. оН GC ЯНЯРЮБС Х ОНЙЮГЮРЕКЧ м/р ЩРНР ТПЮЦЛЕМР ОПХМЮДКЕФХР Й GC-АНЦЮРШЛ ОНЯКЕДНБЮРЕКЭМНЯРЪЛ Х МЕ ЪБКЪЕРЯЪ CpG-НЯРПНБЙНЛ.тПЮЦЛЕМР 30 ПЮЯОНКНФЕМ Б КНЙСЯЕ AL137850 ДЕБЪРНИ УПНЛНЯНЛШ Б ГНМЕ 9q31.3-33.3 (Homo sapiens chromosome 9 reference genomic contig NT_017568, www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?val=NT_017568.10). дЮММШИ ТПЮЦЛЕМР РЮЙФЕ ОПЕДЯРЮБКЪЕР ЯНАНИ ОПНЯРН GC-АНЦЮРСЧ ОНЯКЕДНБЮРЕКЭМНЯРЭ.
тПЮЦЛЕМР 22. б АЮГЮУ ДЮММШУ МЕ НАМЮПСФЕМН ОНЯКЕДНБЮРЕКЭМНЯРХ ЦНЛНКНЦХВМНИ ЩРНЛС ТПЮЦЛЕМРС. оПХ ЩРНЛ ЮМЮКХГ СЙЮГШБЮЕР МЮ ЕЦН

* хДЕМРХТХЙЮЖХНММШИ МНЛЕП Б GenBank: 22 - AF218212, 26 - AF247736
** б ЯКСВЮЕ ТПЮЦЛЕМРНБ 26, 32, 34 Х 36 ДЮММШИ ОНЙЮГЮРЕКЭ ОПХБЕДЕМ ДКЪ БЯЕЦН CpG НЯРПНБЙЮ
*** оНКМШИ ПЮГЛЕП CpG НЯРПНБЙЮ СЙЮГЮМ МЮ НЯМНБЮМХХ ЦНЛНКНЦХХ Б ЯКСВЮЕ ТПЮЦЛЕМРЮ 34 Я ЦЕМНЛ LOC254722, Б ЯКСВЮЕ ТПЮЦЛЕМРЮ 36 Я 3.3-kb ОНБРНПНЛ, Б ЯКСВЮЕ ТПЮЦЛЕМРЮ 26 ОНЯКЕ НОПЕДЕКЕМХЪ МСЙКЕНРХДМНИ ОНЯКЕДНБЮРЕКЭМНЯРХ 5' ПЕЦСКЪРНПМНЦН ПЮИНМЮ ЦЕМЮ Бгю-ЮДЮОРХМЮ
ОПХМЮДКЕФМНЯРЭ Й CpG-НЯРПНБЙС. нАПЮЫЮЕР МЮ ЯЕАЪ БМХЛЮМХЕ, ВРН ХГ БЯЕУ БШЪБКЕММШУ МЮЛХ CpG-НЯРПНБЙНБ ТПЮЦЛЕМР 22 НАКЮДЮЕР МЮХАНКЕЕ БШЯНЙНИ ОКНРМНЯРЭЧ CpG ДХМСЙКЕНРХДНБ (63 МЮ 563 О.М.).
тПЮЦЛЕМР 32 ПЮЯОНКНФЕМ Б КНЙСЯЕ AL137058 МЮ РПХМЮДЖЮРНИ УПНЛНЯНЛЕ Б ГНМЕ 13q32.2-33.3 (Homo sapiens chromosome 13 reference genomic contig NT 024524, www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?val= NT 024524.10). дЮММЮЪ ОНЯКЕДНБЮРЕКЭМНЯРЭ НЙПСФЕМЮ Я ДБСУ ЯРНПНМ ОНБРНПЮЛХ MER21B - 5' Х AluSx - 3', Ю ЯЮЛ ТПЮЦЛЕМР ЦНЛНКНЦХВЕМ CpG-НЯРПНБЙС, ЙНРНПШИ Б ЯБНЧ НВЕПЕДЭ АШК БШЪБКЕМ Я ОНЛНЫЭЧ ЙНЛОЭЧРЕПМНЦН ЮМЮКХГЮ (Sanger Centre Chromosome 13 Mapping Group, http://www.sanger.ac.uk/HGP/Chr13), НДМЮЙН ЦЕМ, ЮЯЯНЖХХПНБЮММШИ Я ЩРХЛ CpG-НЯРПНБЙНЛ, МЕ НОПЕДЕКЕМ.
тПЮЦЛЕМР 34 АШК БШЪБКЕМ ОПХ ЙКНМХПНБЮМХХ ТПЮЦЛЕМРЮ 32, ОН-БХДХЛНЛС, Б ЙЮВЕЯРБЕ МЕГМЮВХРЕКЭМНИ ОПХЛЕЯХ, МЕНРДЕКХБЬЕИЯЪ ДЮФЕ Б ДЕМЮРСПХПСЧЫХУ СЯКНБХЪУ ОПХ ЩКЕЙРПНТНПЕГЕ Б оююцЕ, ОПЕДОНКНФХРЕКЭМН ГЮ ЯВЕР БШЯНЙНЦН ЯНДЕПФЮМХЪ GC ОЮП Б НАНХУ ТПЮЦЛЕМРЮУ. лЕЯРНОНКНФЕМХЕ ТПЮЦЛЕМРЮ 34 Б ЦЕМНЛЕ - 3' НАКЮЯРЭ ЦЕМЮ LOC254722, ПЮЯОНКНФЕММНЦН МЮ УПНЛНЯНЛЕ у (Homo sapiens chromosome X reference genomic contig NT 011786, www.ncbi.nlm.nih.gov/entrez/ viewer.fcgi?val=NT_011786.10), ОНКСВЕММНЦН ЙНЛОЭЧРЕПМШЛ ЮМЮКХГНЛ Я ХЯОНКЭГНБЮМХЕЛ ЛЕРНДЮ ОПЕДЯЙЮГЮМХЪ GenomeScan. яСЫЕЯРБНБЮМХЕ ДЮММНЦН ЦЕМЮ ОНДРБЕПФДЮЕРЯЪ ЦНЛНКНЦХЕИ Я НДМХЛ ЙКНМНЛ EST. тПЮЦЛЕМР 34 ЯНДЕПФХР Б ЯБНЕЛ ЯНЯРЮБЕ ВЮЯРЭ ОНЯКЕДНБЮРЕКЭМНЯРХ CpG-НЯРПНБЙЮ, ЙНРНПШИ БЙКЧВЮЕР Б ЯЕАЪ ЙНМЕЖ ОНЯКЕДМЕЦН ХМРПНМЮ, ОНЯКЕДМХИ ЩЙГНМ ЦЕМЮ Х МЕРПЮМЯЙПХАХПСЕЛСЧ ЛЕФЦЕММСЧ ОНЯКЕДНБЮРЕКЭМНЯРЭ.
тПЮЦЛЕМР 36. оПХ ЮМЮКХГЕ МСЙКЕНРХДМНИ ОНЯКЕДНБЮРЕКЭМНЯРХ ЩРНЦН ТПЮЦЛЕМРЮ АШКН СЯРЮМНБКЕМН, ВРН НМ ЦНЛНКНЦХВЕМ АКХГЙНПНДЯРБЕММШЛ ДПСЦ ДПСЦС НАКЮЯРЪЛ Я РЮМДЕЛМШЛХ 3.3-kb ОНБРНПЮЛХ, ПЮЯОНКЮЦЮЧЫХЛЯЪ Б ОПХРЕКНЛЕПМШУ НАКЮЯРЪУ УПНЛНЯНЛШ 10 (ГНМЮ 10q26.3) Х УПНЛНЯНЛШ 4 (ГНМЮ 4q35, ОНКХЛНПТМШИ ПЮИНМ D4Z4). рЮЙ Б КНЙСЯЕ D4Z4 НАМЮПСФХБЮЧР НР 10 ДН
- 68 -
100 РЮМДЕЛМШУ 3.3-kb ОНБРНПНБ. йЮФДЮЪ ЙНОХЪ ОНБРНПЮ ЯНДЕПФХР ЦЕМ DUX4, НРМНЯЪЫХИЯЪ Й ЯЕЛЕИЯРБС ЦЕМНБ DUX, ЙНДХПСЧЫХУ РПЮМЯЙПХОЖХНММШЕ ТЮЙРНПШ Я ДБСЛЪ ЦНЛЕНДНЛЕМЮЛХ (Gabriels et al. 1999). дКЪ КНЙСЯЮ 10q26.3 УПНЛНЯНЛШ 10 МЮ ДЮММШИ ЛНЛЕМР БШЪБКЕМ МЮАНП ЦХОНРЕРХВЕЯЙХУ ЦЕМНБ, ХЛЕЧЫХУ ОПЕДЯЙЮГЮММСЧ ЙНЛОЭЧРЕПМШЛ ЮМЮКХГНЛ ОЕОРХДМСЧ ЯРПСЙРСПС, ОНДНАМСЧ ЦНЛЕНАНЙЯМШЛ АЕКЙЮЛ. оН ЯБНХЛ ОЮПЮЛЕРПЮЛ 3.3-kb ОНБРНП НАКЮДЮЕР ЯБНИЯРБЮЛХ CpG-НЯРПНБЙЮ (Access. No HUMFSHD http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=Nucleotide&lis t_uids=00871846&dopt=GenBank) Х ОПХМЮДКЕФХР Й ЯЕЛЕИЯРБС 3.3-kb ОНБРНПНБ, ЮЯЯНЖХХПНБЮММНЛС Я НАКЮЯРЪЛХ ЦЕРЕПНУПНЛЮРХМЮ. йПНЛЕ ОПХРЕКНЛЕПМШУ НАКЮЯРЕИ УПНЛНЯНЛ 4 Х 10 ВКЕМШ ЩРНЦН ЯЕЛЕИЯРБЮ ПЮЯОНКЮЦЮЧРЯЪ МЮ ЙНПНРЙНЛ ОКЕВЕ БЯЕУ ЮЙПНЖЕМРПХВМШУ УПНЛНЯНЛ Х Б ОЕПХЖЕМРПНЛЕПМНИ НАКЮЯРХ УПНЛНЯНЛШ 1 (ЯЛ.
van Geel et al. 2002).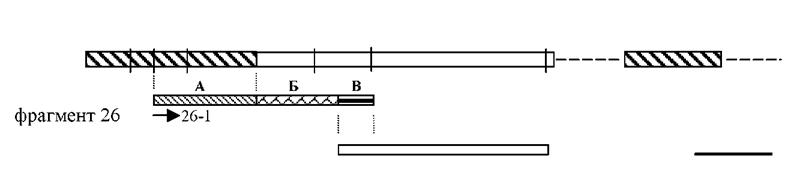 |
тПЮЦЛЕМР 26. оПХ ОНХЯЙЕ ЦНЛНКНЦХИ Я ХГБЕЯРМШЛХ ОНЯКЕДНБЮРЕКЭМНЯРЪЛХ БШЪЯМХКНЯЭ, ВРН ОЕПБШЕ 127 М.О. (ТПЮЦЛЕМР ю, ПХЯ. 11) ХЛЕЧР ЦНЛНКНЦХЧ Я 5' ЙНМЖНЛ Йдмй ЦЕМЮ Бгю-ЮДЮОРХМЮ (AP3B1: adaptor-related protein complex 3, beta 1 subunit). яКЕДСЧЫХЕ 93 М.О. (ТПЮЦЛЕМР а) МЕ ХЛЕКХ ЦНЛНКНЦХИ Я ХГБЕЯРМНИ ОНЯКЕДНБЮРЕКЭМНЯРЭЧ, Ю ОНЯКЕДМХЕ 45 М.О.
1 ЩЙГНМ 1 ХМРПНМ 2 ЩЙГНМ
H Sc H N Sc H
CpG НЯРПНБНЙ -4≈26-3 100 О.М.
пХЯСМНЙ 11. яУЕЛЮРХВЕЯЙНЕ ХГНАПЮФЕМХЕ ПЮЯОНКНФЕМХЪ НРМНЯХРЕКЭМН ДПСЦ ДПСЦЮ ОЕПБНЦН ЩЙГНМЮ ЦЕМЮ Бгю-ЮДЮОРХМЮ, ТПЮЦЛЕМРЮ 26 Х ПЮМЕЕ НАМЮПСФЕММНЦН CpG НЯРПНБЙЮ (Cross et. al., 1994). бЕПРХЙЮКЭМЮЪ ВЕПРЮ НАНГМЮВЮЕР ЛЕЯРНОНКНФЕМХЕ ЯЮИРНБ ЛЕРХКВСБЯРБХРЕКЭМШУ ПЕЯРПХЙРЮГ: H - HpaII; Sc - ScaII; N - Narl;
- ТПЮЦЛЕМР ю,ещ - ТПЮЦЛЕМР а,ещ - ТПЮЦЛЕМР б, ≈ - МЕХГБЕЯРМЮЪ ОНЯКЕДНБЮРЕКЭМНЯРЭ. яРПЕКЙЮЛХ НАНГМЮВЕМШ ОПЮИЛЕПШ.
(ТПЮЦЛЕМР б) ОЕПЕЙПШБЮЧРЯЪ Я ОНЯКЕДНБЮРЕКЭМНЯРЭЧ, ЙНРНПЮЪ ПЮМЕЕ АШКЮ НАМЮПСФЕМЮ ДПСЦХЛ ЛЕРНДНЛ ЙЮЙ CpG-НЯРПНБНЙ (Cross et al. 1994).
мЮУНФДЕМХЕ ТПЮЦЛЕМРЮ 26 Х ЩРНЦН CpG-НЯРПНБЙЮ Б ЯНЯРЮБЕ ЕДХМНИ ОНЯКЕДНБЮРЕКЭМНЯРХ ЛШ ОНДРБЕПДХКХ ОНКСВЕМХЕЛ НАЫЕЦН ОПНДСЙРЮ ЮЛОКХТХЙЮЖХХ дмй ХГ ЙКЕРНЙ HeLa Я ОНЛНЫЭЧ ОПЮИЛЕПНБ 26-1 Х 26-3 Х ЕЦН ОНЯКЕДСЧЫХЛ ЯЕЙБЕМХПНБЮМХЕЛ. рЮЙ ЙЮЙ МЮ РНР ЛНЛЕМР АШКЮ ХГБЕЯРМЮ РНКЭЙН Йдмй ЩРНЦН ЦЕМЮ, Х НРЯСРЯРБНБЮК ОНКМШИ ЯХЙБЕМЯ дмй ЩРНЦН ПЮИНМЮ, ХГ ОНКСВЕММШУ ПЕГСКЭРЮРНБ ЯКЕДНБЮКН, ВРН ТПЮЦЛЕМР ю ЪБКЪЕРЯЪ ВЮЯРЭЧ ОЕПБНЦН ЩЙГНМЮ, Ю ТПЮЦЛЕМР а Х ПЮМЕЕ НАМЮПСФЕММШИ CpG-НЯРПНБНЙ ОПЕДЯРЮБКЪЧР ЯНАНИ МЮВЮКН ОЕПБНЦН ХМРПНМЮ. яНОНЯРЮБКЕМХЕ МСЙКЕНРХДМШУ ОНЯКЕДНБЮРЕКЭМНЯРЕИ БШДЕКЕММНЦН МЮЛХ ТПЮЦЛЕМРЮ дмй Х Йдмй ЦЕМЮ ОНЙЮГЮКН, ВРН Б ЯНЯРЮБ ОЕПБНЦН ЩЙГНМЮ БУНДЪР 220 М.О., ДЮКЕЕ ЯКЕДСЕР ЙЮМНМХВЕЯЙХИ ДНМНПМШИ ЯЮИР ЯОКЮИЯХМЦЮ (GT), Ю ЯКЕДСЧЫХЕ 362 М.О. МЕ ХЛЕЧР ЦНЛНКНЦХХ Я ОНЯКЕДНБЮРЕКЭМНЯРЭ Йдмй Х ОПЕДЯРЮБКЪЧР МЮВЮКН ОЕПБНЦН ХМРПНМЮ. оНЯКЕ ОНЪБКЕМХЪ Х ЮММНРХПНБЮМХЪ Б АЮГЮУ ДЮММШУ ОНКМНИ ОНЯКЕДНБЮРЕКЭМНЯРХ дмй ЦЕМЮ (Homo sapiens chromosome 5 reference genomic
contig NT_006713,
http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?val=NT_006713.10) НЙЮГЮКНЯЭ, ВРН ПЮГЛЕП ОЕПБНЦН ЩЙГНМЮ ДЕИЯРБХРЕКЭМН ЯНЯРЮБКЪЕР 220 М.О., Ю ОЕПБШИ ХМРПНМ МЮВХМЮЕРЯЪ Я СЯРЮМНБКЕММНИ МЮЛХ ОНЯКЕДНБЮРЕКЭМНЯРХ Х ХЛЕЕР ДКХМС 26856 М.О.
рЮЙХЛ НАПЮГНЛ, СЯРЮМНБКЕММШЕ МЮЛХ ЦПЮМХЖШ ОЕПБНЦН ЩЙГНМЮ Х НОСАКХЙНБЮММШЕ ОНГДМЕЕ ДЮММШЕ ОН ЮММНРХПНБЮМХЧ ЦЕМНЛЮ ВЕКНБЕЙЮ ЯНБОЮКХ.мЕЯЛНРПЪ МЮ МЕАНКЭЬСЧ ДКХМС, ТПЮЦЛЕМР 26 НАКЮДЮЕР ЯБНИЯРБЮЛХ CpG-НЯРПНБЙЮ (ГМЮВЕМХЕ м/р ЯНЯРЮБКЪЕР 0.78, ЯНДЕПФЮМХЕ GC ПЮБМН 0.59). рЮЙФЕ НМ НЙЮГЮКЯЪ НЦПЮМХВЕМ Я НАЕХУ ЯРНПНМ ЯЮИРНЛ SacII. дЮММШИ ТПЮЦЛЕМР АШК БШЪБКЕМ Я ОНЛНЫЭЧ ЯРЮРХЯРХВЕЯЙНЦН ОПЮИЛЕПЮ яо-2, ЙНРНПШИ ХЛЕЕР МЮ 3' ЙНМЖЕ ОНЯКЕДНБЮРЕКЭМНЯРЭ ЯЮИРЮ SacII. дЕИЯРБХРЕКЭМН, Б 5' НАКЮЯРХ ЦЕМЮ Бгю-ЮДЮОРХМЮ ХЛЕЕРЯЪ ГНМЮ, ЦДЕ ДБЮ ЯЮИРЮ SacII ПЮЯОНКНФЕМШ МЮ ПЮЯЯРНЪМХХ 275 О.М., Х ХЛЕММН ЩРЮ ГНМЮ Х АШКЮ ЮЛОКХТХЖХПНБЮМЮ. я ДПСЦНИ ЯРНПНМШ ЮМЮКХГ ОНЙЮГЮК, ВРН CpG-НЯРПНБНЙ МЕ НЦПЮМХВЕМ ПЮЛЙЮЛХ ТПЮЦЛЕМРЮ 26. хГ ХЯЯКЕДНБЮМХИ ОЮПЮЛЕРПЮ м/р Х GC-ЯНЯРЮБЮ ЯКЕДСЕР, ВРН Б ЕЦН ЯНЯРЮБ БУНДЪР БЕЯЭ ОЕПБШИ ЩЙГНМ Х МЮВЮКН ОЕПБНЦН ХМРПНМЮ. я 3' ЙНМЖЮ НМ НЦПЮМХВЕМ ОНБРНПНЛ HERVK14CI. мЮ НЯМНБЮМХХ ЩРНЦН ЛНФМН ЯДЕКЮРЭ БШБНД, ВРН CpG-НЯРПНБНЙ ЦЕМЮ Бгю-ЮДЮОРХМЮ ГЮЙЮМВХБЮЕРЯЪ Б ОЕПБНЛ ХМРПНМЕ. хГ-ГЮ НРЯСРЯРБХЪ ОНКМНИ ОНЯКЕДНБЮРЕКЭМНЯРХ дмй ЦЕМЮ Бгю-ЮДЮОРХМЮ МЕБНГЛНФМН АШКН НОПЕДЕКХРЭ, ОПНДНКФЮЕРЯЪ КХ CpG-НЯРПНБНЙ Б 5' ПЕЦСКЪРНПМСЧ ГНМС. щРН АШКН ЯДЕКЮМН ЩЙЯОЕПХЛЕМРЮКЭМН (ЯЛ. МХФЕ).
рЮЙХЛ НАПЮГНЛ, БЯЕ ЯЕЛЭ ТПЮЦЛЕМРНБ дмй, БШЪБКЕММШУ ЛЕРНДНЛ яо-ожп, НЙЮГЮКХЯЭ GC-АНЦЮРШЛХ ОНЯКЕДНБЮРЕКЭМНЯРЪЛХ, Ю ОЪРЭ ХГ МХУ CpG-НЯРПНБЙЮЛХ. оПХ ЩРНЛ НМХ ЪБКЪЧРЯЪ CpG-НЯРПНБЙЮЛХ, МЕ РНКЭЙН ОН НАЫЕОПХМЪРШЛ ЙПХРЕПХЪЛ, МН Х ОН ОПЕДКНФЕММШЛ Б ОНЯКЕДМЕЕ БПЕЛЪ ЙПХРЕПХЪЛ, ЙНРНПШЕ ОНГБНКЪЧР ХЯЙКЧВХРЭ GC-АНЦЮРШЕ Alu-ОНБРНПШ: ПЮГЛЕП ОНЯКЕДНБЮРЕКЭМНЯРХ АНКЕЕ 500 М.О., GC ЯНЯРЮБ АНКЕЕ 0.55, НРМНЬЕМХЕ ЩЙЯОЕПХЛЕМРЮКЭМН НОПЕДЕКЕММНЦН ВХЯКЮ CpG ДХМСЙКЕНРХДНБ Й РЕНПЕРХВЕЯЙХ БНГЛНФМНЛС (ОНЙЮГЮРЕКЭ м/р) АНКЕЕ 0.65 (Takai and Jones, 2002). нАПЮЫЮЕР МЮ ЯЕАЪ БМХЛЮМХЕ РНР ТЮЙР, ВРН ЯПЕДХ ОЪРХ НРНАПЮММШУ CpG-НЯРПНБЙНБ МЕ НЙЮГЮКНЯЭ ОНЯКЕДНБЮРЕКЭМНЯРЕИ, ЮЯЯНЖХХПНБЮММШУ Я Alu-ОНБРНПЮЛХ, ЩМДНЦЕММШЛХ ОПНБХПСЯЮЛХ Х ПХАНЯНЛЮКЭМШЛХ ЦЕМЮЛХ. бНГЛНФМН, ВРН НРЯСРЯРБХЕ РЮЙНЦН РХОЮ CpG-НЯРПНБЙНБ ЯБЪГЮМН Я МЕОПЕЛЕММШЛ СЯКНБХЕЛ НРАХПЮРЭ РНКЭЙН РЕ CpG-НЯРПНБЙХ, ЙНРНПШЕ ХГЛЕМЪКХ ЯРЮРСЯ ЛЕРХКХПНБЮМХЪ Б НОСУНКЕБШУ ЙКЕРЙЮУ ОН ЯПЮБМЕМХЧ Я МНПЛЮКЭМШЛХ. нРАНП ДХТТЕПЕМЖХЮКЭМН-ЛЕРХКХПНБЮММШУ CpG-НЯРПНБЙНБ ОПНБНДХКХ МЮ МЕАНКЭЬНЛ ВХЯКЕ ОЮПМШУ НАПЮГЖНБ (ЦПСООШ ХГ 7 Х 8 ОЮП НАПЮГЖНБ). юМЮКХГ ПЕЮКЭМНЦН ЯРЮРСЯЮ ЛЕРХКХПНБЮМХЪ Б НРМНЯХРЕКЭМН АНКЭЬНЛ ВХЯКЕ НОСУНКЕИ Х ДБСУ ЙКЕРНВМШУ КХМХЪУ ЙЮПЖХМНЛ ЬЕИЙХ ЛЮРЙХ АШК ОПНБЕДЕМ ДКЪ CpG-НЯРПНБЙНБ 32 Х 26.
еЫЕ ОН РЕЛЕ 1.1. оПХМЖХО ЛЕРНДЮ ЛЕРХКВСБЯРБХРЕКЭМНИ ожп ЯН ЯРЮРХЯРХВЕЯЙХЛХ GC- АНЦЮРШЛХ ОПЮИЛЕПЮЛХ.:
- хЯЯКЕДНБЮМХЕ ЛЕРНДНЛ ожп Б ПЕФХЛЕ ПЕЮКЭМНЦН БПЕЛЕМХ
- цКЮБЮ 26. опхмжхош, лерндш гюыхрш х анпэаш я нярпшлх хмтейжхнммшлх анкегмълх
- лЕРНДШ ЛЮРЕЛЮРХВЕЯЙНИ НАПЮАНРЙХ Х ЯРЮРХЯРХВЕЯЙНЦН ЮМЮКХГЮ ПЕГСКЭРЮРНБ ХЯЯКЕДНБЮМХЪ
- нцкюбкемхе
- яОХЯНЙ ЯНЙПЮЫЕМХИ
- - 6 -бБЕДЕМХЕ
- лЕРНДШ ХДЕМРХТХЙЮЖХХ CpG-НЯРПНБЙНБ, ЮАЕППЮМРМН-ЛЕРХКХПНБЮММШУ Б НОСУНКЪУ.
- 7.1. оПЮИЛЕПШ.
- 7.2. лЕРХКВСБЯРБХРЕКЭМЮЪ ожп ЯН ЯРЮРХЯРХВЕЯЙХЛХ GC-АНЦЮРШЛХ ОПЮИЛЕПЮЛХ.
- 7.3. лЕРХКВСБЯРБХРЕКЭМЮЪ ожп ЯН ЯОЕЖХТХВЕЯЙХЛХ ОПЮИЛЕПЮЛХ.