ДИНАМИКА ВЕСОВЫХ ИЗМЕНЕНИЙ НА СТАДИИ ПИТАНИЯ
Для характеристики весовых изменений на стадии питания мы ввели специальные показатели: абсолютное увеличение веса и удельный привес (Балашов, 19586). Абсолютное увеличение веса по отношению к голодной стадии для любого дня питания дается в процентах и вычисляется по формуле
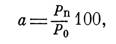
где а — абсолютное увеличение веса для n-го дня кровососания, Рп — вес в п-й день питания и Р0 — вес голодного клеща в миллиграммах.
где d — удельный привес за сутки, Рп+1 и Рп — вес за последовательные дни питания и t — время между двумя измерениями в сутках.
При уменьшении веса клеща по сравнению с исходным величина удельного привеса меньше 100%-, а при увеличении веса — больше 100%.Абсолютное увеличение веса и удельный привес отчасти сходны с абсолютной и удельной скоростями роста. Однако они включают в себя как изменения в весе за счет роста клеща, так и простое накопление крови в дивертикулах кишечника.
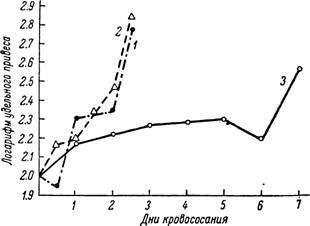
Рис. 165. Изменения величины удельного привеса в период питания у Ixodes ricinus.
Удельный привес до некоторой степени соответствует средней скорости кровососания.
і — личинки; 2 — нимфы; з — самки.
Величина удельного привеса характеризует скорость изменений веса в процентах за любые дни кровососания и вычисляется по формуле
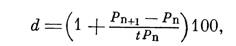
Неравномерность скорости увеличения размеров и веса тела иксодовых клещей во время питания была впервые обнаружена А. Лисом (Lees, 1952) у самок Ixodes ricinus. В первые 5—6 суток питания вес их тела увеличивался довольно равномерно и достигал 50 мг, тогда как за последние сутки кровососания их вес сразу же увеличивался до 200 мг. Универсальность этой особенности питания была в дальнейшем подтверждена не только на самках, но и на неполовозрелых фазах развития представителей большинства родов мировой фауны (Балашов, 1956а, 19586; Хейсин и Лаврененко, 1956; Kitaoka a.
Yajima, 1958а). В то же время на основании полученных нами данных мы считаем, что стадию питания по характеру изменений в весе и размерах и особенно в связи с морфо-физиологическими изменениями в организме следует подразделять не на два, а на три периода.Первые 6—24 часа с момента прикрепления первоначальный вес клеща уменьшается или же остается без заметных изменений. Особенно значительная потеря веса в первые сутки прикрепления наблюдается у личинок и нимф. Обычно она не превышает 10—20%, но в исключительных случаях, как например у личинок Hyalomma anatolicum, достигает почти 30—40% от исходного веса. У личинок и нимф Ixodes ricinus этот период заметно укорочен, а иногда полностью отсутствует (рис. 165), чем, вероятно, и объясняется игнорирование его Лисом (Lees, 1952). У Hyalomma asiaticum и Haemaphysalis punctata он может растягиваться на 2—3 суток (рис. 166). У самок первоначальное уменьшение веса незначительно, хотя они иногда не питаются несколько дней после прикрепления.
Уменьшение веса в первые часы или даже сутки после прикрепления связано с интенсивной потерей воды за счет роста испарения при температуре прикожного слоя воздуха в 26—36°. При кормлении клещей на рептилиях при температуре не выше 25—30° уменьшение веса в первые дни прикрепления вообще может отсутствовать или бывает очень незначительным. Вероятно, переход клещей от свободного существования к паразитированию на теле хозяина наряду с другими перестройками в организме нарушает водонепроницаемость покровов тела. В пользу этого
косвенно свидетельствует малая устойчивость к высыханию у недопитавшихся клещей после их удаления с хозяина.
Отсутствие питания или очень медленное поступление пищи в кишечник сразу после прикрепления связано с некоторой недоразвитостью органов пищеварения у голодных особей. Клещам требуется некоторое время для морфо-физиологических перестроек в их организме, лишь после этого они становятся способными к нормальному питанию. Длительность этого периода, названного нами подготовительным, определяется видовыми особенностями клещей, фазой развития, физиологическим состоянием прикрепляющихся особей и характером воздействия факторов внешней среды.
Быстрее всего он заканчивается у клещей 2—3-й степеней истощения,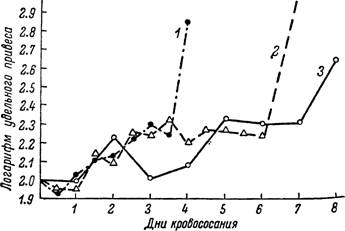
Рис. 166. Изменения величины удельного привеса в период питания у Hyalomma asiaticum.
проявляющих значите л ь- ^ ную агрессивность.
і — личинки; 2 — нимфы; 3 — самки.
Собственно питание клещей на теле хозяина по скорости принятия пищи и физиологическим процессам в организме подразделяется на два периода. Большую часть времени кровососания занимает период роста. В это время происходит быстрое переваривание крови хозяина и за счет ее ассимиляции интенсивный рост покровов и внутренних органов. Ветви кишечника, несмотря на значительное общее увеличение веса, растянуты умеренно, сохраняют многочисленные перетяжки и имеют морщинистую наружность. Окончанием этого периода можно считать начало быстрого растяжения тела клеща поступающей кровью за 0.5—1.5 суток до отпадения.
У самок на этом этапе в первые сутки происходит быстрое возрастание величины удельного привеса, а затем эта величина 4—6 дней держится на достигнутом уровне, колеблясь в пределах 170—220%. Примером подобного идеального случая могут служить самки Ixodes ricinus (рис. 165). Последнее также отмечено для самок Boophilus caudatus (Kitaoka a. Yajima, 1958а), у которых выделяются три периода роста: от 3 до 10 мг, от 10 до 80 мг и от 80 до 450 мг (рис. 167), причем первый период занимает большую часть времени питания. Исследования других видов иксодид и особенно питания личинок и нимф, включая и данные указанных выше авторов по личинкам Haemaphysalis bispinosa, не подтверждают постоянства существования двух периодов в росте иксодовых клещей до начала периода растяжения (3-я стадия в работах японских исследователей). У самок некоторых видов в период роста часто наблюдаются заметные подъемы и падения скорости кровососания. Наблюдавшееся в наших опытах закономерное падение величины удельного привеса до 110—130% и последующий резкий подъем до 300—350%, вероятно, были связаны с торможением кровососания у неоплодотворенных самок и быстрым ее возрастанием после оплодотворения. Особенно типично в этом отношении питание половозрелых Hyalomma asiaticum.
У неоплодотворенных самок этого вида задержка питания начинается уже на 4-е сутки прикрепления. Самцы же становятся способными к оплодотворению лишь на 5-е сутки. В связи с этой особенностью при одновременной посадке самцов и самок на хозяине у последних всегда наблюдается кратковременное замедление кровососания (рис. 166).Незначительное уменьшение величины удельного привеса, отмеченное для большинства видов перед началом быстрого насыщения клеща в последние 0.5—1 сутки кровососания, вероятно, связано с определенной физиологической перестройкой в организме перед сильным растяжением покровов идиосомы.
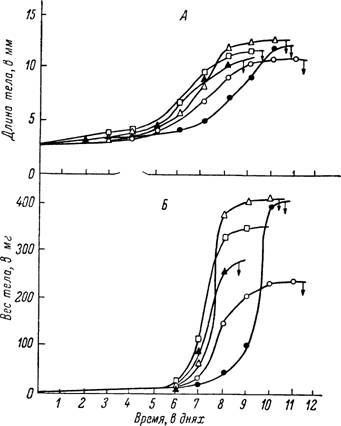
Рис. 167. Изменения длины (А) и веса (Б) тела у самок Boophilus caudatus во время питания. Стрелки обозначают время отпадения напитавшихся особей (по Kitaoka a. Yajima, 1958а).
Изменения величины удельного привеса у нимф имеют ряд своих особенностей. У Dermacentor pictus, Z). marginatus, Hyalomma asiaticum и H. anatolicum, обладающих длительным нимфальным питанием и значительным увеличением веса, изменения величины удельного привеса во многом сходны с таковыми у самок (рис. 166). У них хорошо выражен 2—4-дневный период с относительно постоянным удельным привесом в 150—250%. Наоборот, у Ixodes ricinus, Haemaphy salts punctata, Rhipicephalus turani- cus и Rh. bursa с коротким периодом питания и сравнительно небольшим увеличением веса все время происходит постоянное нарастание удельного привеса (рис. 165).
Для личинок с их коротким питанием и еще меньшим увеличением в весе постоянный рост величины удельного привеса представляет уже общую отличительную черту. Однако и у личинок, и у нимф после первоначально быстрого подъема скорости кровососания всегда существует продолжительный период с более медленным ее увеличением.
При кровососании у личинок и нимф наблюдаются значительно большие индивидуальные колебания в удельном привесе по сравнению с самками. Возможно, что меньшие размеры прикрепительного аппарата не позволяют им достаточно глубоко внедриться в покровы хозяина, а это ставит их в большую зависимость от различных местных реакций в области прикрепления.
Таким образом, для питания личинок и нимф мелких видов иксодовых клещей характерен период роста с относительно постоянным умеренным увеличением удельного привеса, а для самок и нимф крупных видов — постоянство величины удельного привеса в этот период.
У самок период роста занимает 5—7, у нимф 2—4 и у личинок 1.5—3 суток. За это время вес самок увеличивается в 10—20, нимф в 5—20 и личинок в 3—5 раз, что составляет от Ѵв до Ѵ4 веса полностью напитавшегося клеща.За счет непрерывного переваривания крови хозяина происходят интенсивные процессы роста, и к концу этого периода сильно увеличившийся в размерах клещ приобретает способность к быстрому поглощению огромной массы крови, обеспечивающей его дальнейшее развитие по отпадении.
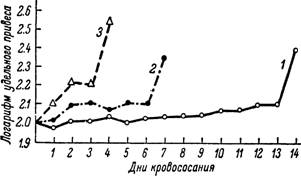
Рис. 168. Изменения величины удельного привеса в период кровососания у личинок Haemaphysalis sulcata, питавшихся на зеленой ящерице (Lacerta viridis) при различной температуре.
После продолжительного периода с относительно постоянным приростом в весе за 12—24 часа до отпадения начинается необычайно быстрое увеличение размеров клеща. К концу его, перед отпадением, клещ буквально раздувается от поглощенной крови хозяина. За последние сутки питания клещи увеличиваются в весе больше, чем за весь предшествующий многодневный период. В дальнейшем мы будем называть его за столь быстрое увеличение размеров периодом растяжения. Вес самок в эти последние часы увеличивается у мелких видов с 60—100 до 200—600 мг и у крупных с 150—200 до 600—1200 мг (табл. 23). Нимфы крупных видов увеличиваются с 2—4 до 10—30 мг и мелких — с 0.5—1 до 3—5 мг; личинки большинства изученных видов — с 0.1—0.2 до 0.3—0.6 мг.
Величина удельного привеса достигает 400—700% против 150—200% в предшествующий период. У самок последний этап кровососани я обычно растянут несколько дольше и соответственно удельный привес меньше, чем у личинок и нимф.
I — 20°; 2 — 25°; 3 — 30°.
Интересно, что у самок Ixodes persulcatus и /. ricinus окончание периода насыщения обычно тесно связано с суточным ритмом отпадения и происходит во время нахождения коровы в хлеву на покое. При пастьбе лишь немногие клещи заканчивают насыщение. Вероятно, энергичные движения животного на пастбище, способствуя отпадению насосавшихся клещей, в то же время до некоторой степени препятствуют окончанию кровососания.
Последний период кровососания иксодовых клещей можно с известными основаниями сравнивать со всем актом питания других кровососв.
На первое место теперь выступают процессы растяжения покровов идиосомы и стенок кишечника. Клещ поглощает гигантские количества крови по сравнению с его первоначальным весом. Фактически, однако, растяжение покровов и кишечника не выходит в это время из рамок, свойственных другим кровососущим членистоногим, так как растяжению подвергается организм клеща, успевший до этого значительно увеличиться в размерах. Вес клеща по сравнению с достигнутым к концу предшествующего этапа увеличивается не больше чем в 4—10 раз. Это довольно близко к увеличению веса у насосавшихся орнитодорин, не имеющих особого периода роста.При кормлении на холоднокровных животных соотношение в продолжительности отдельных периодов кровососания в известных границах определяется температурой. У личинок и нимф Haemaphysalis sulcata, питавшихся на зеленых ящерицах при 20, 25 и 30°, наблюдалось заметное удлинение первых двух периодов при понижении температуры. Пропорционально удлинению периода роста происходило уменьшение величины удельного привеса, тогда как продолжительность периода растяжения менялась в значительно меньшей степени (рис. 168). Сходные данные были получены при кормлении личинок и нимф Ixodes ricinus на прытких ящерицах и кавказских агамах.
Еще по теме ДИНАМИКА ВЕСОВЫХ ИЗМЕНЕНИЙ НА СТАДИИ ПИТАНИЯ:
- Динамика изменений интервала Q-T и его дисперсии в зависимости от стадии и клинико-функциональных особенностей ИМ
- Стадии изменений
- 13.3 Динамика: долгосрочные изменения
- МЕРЫ ВЕСОВ И ОБЪЕМОВ
- Изменения парадоксального сна на доклинической и клинической стадии у животных и при болезни Паркинсона у человека
- Динамика изменения суточного профиля АД в зависимости от натрийуреза
- Возрастная динамика изменений эндотелия визуально неизмененных сосудов
- Изменение состояния пульпы у обследованных групп пациентов в динамике
- Динамика изменения морфологических показателей слизистой оболочки желудка в различные сроки после эрадикационной терапии
- МЕРЫ ОБЪЕМА И ВЕСОВ ИЗ СБОРНИКА ЙУХАННЬІ ИБН САРАФИЙУНА?
- Изменения частоты факторов у больных раком желудка и толстой кишки при I-II стадии распространенности процесса.
- Изменения частоты факторов у больных раком желудка и толстой кишки при III-IV стадии распространенности процесса.
- 32. Типичные стадии в динамике работоспособности субъекта труда в течение рабочей смены. Кривая работы. Продуктивность труда и работоспособность.
- Динамика изменений замедленной желудочковой активности у больных с острым ИМ и в постинфарктном периоде в зависимости от наличия и степени выраженности сердечной недостаточности
- Динамика изменений изучаемых показателей метаболического гомеостаза у пациенток с миомой матки при включении эмоксипина в общепринятый метод терапии
- Особенности организации питания, медицинский контроль и гигиенические требования к рациону питания детей и подростков в образовательных учреждениях
- Организация диетического питания и питания по лечебному пайку
- Часть II. Неправильное питание и его последствия Заболевания, связанные с употреблением в пищу недоброкачественных продуктов питания
- Изменение реактивности при нарушении кислотно-основного равновесия (по изменению частоты сердечных сокращений)
- Стадии лихорадки.