окислительная модификация биологических структур
должна, казалось бы, начинаться уже с момента рождения животного организма. Однако, на наш взгляд, следует говорить не об изначальности старения, постулируемой в большинстве теорий, а о наличии объективных природных факторов (гипероксии, радиации, тепла и др.), которые наряду с позитивной ролью в некоторых адаптировавшихся к ним про-цессах оказывают непрерывное «давление» и противодействие реализации программы развития организма с момента его рождения.
Сначала это действие со стороны, скажем, дисбаланса D (ПО – АО) незначительно ввиду малости его значения, и все ткани, органы и системы регуляции растут и функционируют относительно нормально в соответствии с программой развития, заложенной природой. Однако со временем во всех клетках и тканях организма величина D (ПО – АО) по разным причинам постепенно возрастает (см. выше), оказывая уже более значимое и, в конечном счёте, необратимое цитотоксическое действие на все биологические структуры.В данной постановке вопроса логичнее было бы считать, что в начальный период развития старение как таковое ещё отсутствует, и негативная составляющая указанных выше физических факторов, своего рода «помеха», достаточно успешно нейтрализуется пока что не дестабилизированными на этом этапе системами защиты (антиоксидантной, микросомальной, репарационной, иммунологической и др.). Действительно к моменту рождения организма активность антиоксидантных ферментов, как, например, у крыс, максимальна (Дубинина, 1989), и это обеспечивает новорожденным животным высокую устойчивость к токсическому действию 02 и его радикалов. В таких защищённых условиях у них поддерживается высокая активность «пролиферативных» ферментов (РНК- и ДНК-полимераз, ДНК-метилтрансферазы и др.), с чем, очевидно, связана максимальная интенсивность пролиферации клеток большинства органов и тканей в пренатальный период и на первом этапе постнатального онтогенеза (Srivastava et al., 1993; Mizuuchi et al., 1994 - cм.
Романенко и др., 1998). При такой интерпретации событий фактически отпадает замечание Аршавского (1995) о том, что «если бы развитие организмов началось со старения, то не только филогенетическая эволюция была бы невозможна, но и сама жизнь».Таким образом, существенно то, что в последующие месяцы и годы жизни ввиду постоянного присутствия фактора D (ПО – АО) отдельные звенья защитных систем организма, в том числе антиоксидантной, постепенно истощаются, а указанный дисбаланс, как уже отмечалось, также постепенно возрастает от некоторого начального (базового) его значения DБ (ПО – АО) до, в конечном счёте, «цитолизного» DЦ (ПО – АО), проходя через промежуточные ведущие к старению значения DС (ПО – АО). В результате с возрастом интенсивность ПОЛ и окислительная модификация различных макромолекул в клетках усиливаются, приводя ко многим негативным последствиям. Одним из примеров, подтверждающих указанные положения, служат результаты определения у дол-гожителей (возраст 100–105 лет) антиоксидантной активности в общей сыворотке крови и в депротеинизированной её фракции путём измерения хемилюминесценции. У долгожителей эта активность по сравнению с таковой у лиц в возрасте от 18 лет до 91 года значительно снижена, свидетельствуя о зависимости этого показателя от возраста. Наличие такого дисбаланса как фактора, инициирующего окислительный стресс, признаётся характерным для процесса старения (Pinzani et al., 1996). Снижение активности антиоксидантных соеди-инений ферментативной и неферментативной природы обнаруживается также в большинстве тканей стареющих животных (Tian et al., 1998; Паранич, 1999).
Постулируемый нами возрастзависимый характер изменения параметра D (ПО – АО) не совпадает точно с указываемым в отдельных работах. Например, в одной из статей авторы (Costantini et al., 1995), подтверждая «системный оксидантный стресс» при старении, отмечают, что у лиц пожилого возраста (80-90 лет) формируется (?) антиоксидантно–прооксидантный дисбаланс. В действительности же такой дисбаланс определяется значительно раньше (см.
выше). Со временем он просто возрастает, активно участвуя в развитии процесса старения на всех его этапах. Сведения о повышении, в частности, уровней АФК и ПОЛ при старении и связанных с ним патологиях достаточно полно отражены в ряде обзоров последних лет (см. Benzi, Moretti, 1995; Gutteridge, 1995; Knight, 1995; Гусев, Панченко, 1997; Ozawa, 1997; Papa, Skulachev, 1997; Johnson et al., 1999; Allen, Tresini, 2000; Knapowski, 2000 и др.).По некоторым данным, динамика изменения соотношения про- и антиоксидантной активностей в ходе онтогенеза может отличаться от указанного выше, особенно в начальный его период. Так, используя метод оценки интенсивности окислительной модификации белков в тканях, установили, что уровень такой модификации у новорожденных детей выше, чем у взрослых (Хамiд, 1999). В первые дни жизни новорожденные очень чувствительны к окислительному повреждению. Защитный эффект в этот период в какой-то мере оказывает рост в организме уровня антиокстданта билирубина (Bottini et al., 2000). Ткани скелетных мышц, сердца, лёгких и семенников 1-дневных поросят характеризуются высоким содержанием продуктов ПОЛ (MDA, гидроперекисей липидов) и высокой активностью SOD при низкой активности глутатионпероксидазы (GPX) и глутатионредуктазы. Однако в период с 1- до 10-дневного возраста содержание продуктов ПОЛ в указанных тканях уменьшается (Снiтинський и др., 1998). Эти данные показывают, что в первые часы и дни после рождения организм новорожденных попадает, по сути, скачкообразно под возмущающее воздействие новой, теперь воздушной среды. Поэтому, несмотря на относительно высокую у них эффективность антиоксидантной системы (см. выше), в самом раннем постнатальном периоде отмечается усиление окислительной модификации липидов и белков. Несколько позже оно ослабляетя в связи с быстрой адаптацией организма новорожденных к гипероксическим условиям земной атмосферы.
К числу таких адаптивных процессов можно отнести, очевидно, увели-чение содержания и активности митохондриальных цитохромов в начальный постнатальный период, показанное в некоторых работах, например, при изучении постнатального онтогенеза энергетического обмена в тканях головного мозга крыс (Kalous et al., 2001).
Как оказалось, активность большинства цитохромов возрастала в период от 5 до 30 дней, в дальнейшем к 6 мес. она постепенно уменьшалась и в период 6-12 мес. не изменялась. Содержание цитохромов увеличивалось от низких оценок у 5-дневных крыс до высоких величин в возрасте одного месаца.Барабой и соавт. (1988), исследуя возрастную кинетику спонтанной хемилюминесценции сыворотки крови у лабораторных крыс и у здоровых людей, установили: «У крыс-самцов наиболее низкие стабильные значения свечения отмечены в возрасте от 2–3 до 18–19 мес.; у более молодых и более старых животных интенсивность хемилюминесценции существенно выше, снижаясь от 1 к 2–3 мес. и вновь возрастая от 18–19 к 24–34 мес. Аналогичная закономерность выявлена и у доноров: минимальные значения свечения отмечены у мужчин и женщин 31–40 лет; в более молодом и более пожилом возрасте интенсивность спонтанной хемилюминесции сыворотки крови существенно выше». Значит, повышенные уровни пероксидации и, следовательно, D (ПО – АО) в крови характерны как для молодого растущего, так и стареющего организмов. Между прочим, с такого рода закономерностью хорошо коррелирует вы-раженная возрастная зависимость некоторых заболеваний, особенно лейкоза. Последний имеет 2 диапазона такой зависимости: «ранний – до 35 лет, когда смертность после быстрого подъёма опять снижается, и поздний – когда смертность с возрастом растёт экспоненциально» (Мамаев и др., 1994). По разным поводам феномен бимодального распределения частоты спонтанного лейкоза и биоэнергетические его аспекты обсуждаются нами и в других разделах (см. п. 1.6.2, 2.5 и 3.6.6).
Таким образом, в живых организмах действует эволюционно заложенный кислородно-перекисный механизм не очень глубоких повреждений, обусловленный появлением и возрастанием в атмосфере Земли свободного О2. Данный причинный фактор представляется первичным, главным и определяющим старение как биологическое явление. От значений дисбаланса D (ПО – АО), отражающих в основном пероксидативную часть метаболизма и обмена веществ, зависит количество и качество действующих в клетках и тканях повреждающих пероксигеназных факторов, устойчивость к ним структур организма и, следовательно, темп его разрушения, старения.
(Gutteridge, 1995). В этом смысле, действительно, генетически запрограммировано не старение, а тип обмена веществ (Фролькис, 1986).Другие факторы окружающей среды усугубляют пероксидативное состояние в клетках и тканях, усиливая процесс старения. Это касается, прежде всего, интенсивной естественной радиации (солнечного излучения) и, особенно, вызванной искусственно. Негативное воздействие их в присутствии О2 в организме может существенно возрастать, поскольку в этом случае проявляется известный в радиобиологии кислородный эффект. А возникновение и развитие в течение последних нескольких веков промышленности, особенно нефтехимической, металлургической, энергетической и ракетно-ядерной, резко ухудшило среду обитания животного и растительного мира, значительно увеличило в ней содержание токсических и канцерогенных веществ с прооксидантными свойствами, что также должно отразиться на старении и проявлении возрастных патологий. В частности, на основании исторического и географического анализа сделан вывод о прямой связи распространения раковых заболеваний с разви-тием общества и его индустриализацией (Goldsmith, 1998).
Кстати, высокие окислительные свойства земной атмосферы и другие факторы окружающей среды вызывают и в неживой природе многие нежелательные явления, также называемые процессами старения. Например, в химии это процессы старения полимеров и других материалов, порча пищевых продуктов и лекарственных препаратов (Эмануэль, 1982).
Для последующего изложения кислородно-перекисной концепции старения и канцерогенеза остановимся ещё на наших представлениях о возможности установления в клетке различных уровней D (ПО – АО), с каждым из которых связываются принципиально разные её состояния. Как отмечалось выше, при низких значениях D (ПО – АО) протекают нормальные синтетические процессы, обеспечивающие дифференцировку клеток и выполнение ими специальных функций. Значения D (ПО – АО), соответствующие этому нормальному невозбуждённому (интерфазному) состоянию, включая временный выход из цикла (состояние покоя) обозначим через DИ (ПО – АО).
Иная ситуация складывается в стимулированной к пролиферации клетке, в которой существенно изменяются уровень и характер протекания биоэнергетических процессов. Так, согласно уже давним сведениям, в период деления клетки происходит резкое ослабление дыхания и потребление ничтожного количества О2 (Rose et al., 1981), а перед делением, во время митоза и вообще при митогенной стимуляции уменьшается содержание АТР (Мэзия, 1963; Lynne et al., 1981).Приведённые факты соответствуют тому, что интенсивность митохон-дриального дыхания во время роста отдельных органов, тканей и организма в целом значительно снижается (Озернюк, 1978). Падение уровня АТР и соотношения ATPADP при переходе клеток из фазы G1 в фазу S служит, как считают (Дмитренко, 1984), необходимым регуляторным моментом для начала и облегчения репликации ДНК, поскольку известно, что, например, повышение уровня АТР в ядерном компартменте угнетает репликацию ДНК и клеточную пролиферацию (Rapoport E. et al., 1979). Кстати, иммортализованные клетки, как показано на примере фибробластов человека (линия KMST-6) при воздействии ATP (0.4 ммоль/л), тоже заметно снижают скорость пролиферации, а синтез их ДНК супрессируется (Li J.-W). С этими материалами согласуются также известные данные о низком содержании митохондрий в эмбриональных и стволовых клетках (см. Von Wagenheim, Peterson, 1998), которые как раз и характеризуются повышенной пролиферативной активностью.
Кроме того, в периодах S и М клеточного цикла снижается концентрация циклического аденозинмонофосфата (сАМР), а уровень cGMP, напротив, возрастает (см. Лю, Ефимов, 1978). Включаются механизмы регуляции деления с участием О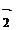 и других АФК (Оberley et al., 1980; Jang et al., 1987; Burdon, 1992, 1995; McCord, 1995; Burdon et al., 1996; Bhunia et al., 1997; Maher, Schubert, 2000), в том числе липо- и циклооксигеназный пути метаболизма арахидоновой кислоты, причём ловушки свободных радикалов, SОD, каталаза, ингибиторы липооксигеназы и гуанилатциклазы отменяют митогенез (Burdon, 1992; Camberini, Leite,1997 др.). А недавно необходимость АФК для фазы синтеза ДНК подтверждена на примере эбриональных фибробластов крысы и полученных из них введением разных пар комплементирующих онкогенов трансформантов. Удаление ростовых факторов (сыворотки) из среды культивирования коррелировало со снижением концентрации АФК в клетках, а при обратном введении сывортки – с повышением внутриклеточного уровня АФК, который максимален был при переходе клеток в S-фазу (Гамалей и др., 2001). Кстати, лактоферрин человека, как Fe-связывающий гликопротеин и ингибитор свободнорадикальных процессов, тоже подавляет пролиферацию клеток, блокирует их клеточный цикл (Damiens et al., 1998). Индуцировать пролиферацию клеток в эксперименте можно, умеренно повышая тем или иным способом образование перекисей липидов, например, скармливанием подопытным животным в избытке того же железа (Whittaker et al., 1992).
и других АФК (Оberley et al., 1980; Jang et al., 1987; Burdon, 1992, 1995; McCord, 1995; Burdon et al., 1996; Bhunia et al., 1997; Maher, Schubert, 2000), в том числе липо- и циклооксигеназный пути метаболизма арахидоновой кислоты, причём ловушки свободных радикалов, SОD, каталаза, ингибиторы липооксигеназы и гуанилатциклазы отменяют митогенез (Burdon, 1992; Camberini, Leite,1997 др.). А недавно необходимость АФК для фазы синтеза ДНК подтверждена на примере эбриональных фибробластов крысы и полученных из них введением разных пар комплементирующих онкогенов трансформантов. Удаление ростовых факторов (сыворотки) из среды культивирования коррелировало со снижением концентрации АФК в клетках, а при обратном введении сывортки – с повышением внутриклеточного уровня АФК, который максимален был при переходе клеток в S-фазу (Гамалей и др., 2001). Кстати, лактоферрин человека, как Fe-связывающий гликопротеин и ингибитор свободнорадикальных процессов, тоже подавляет пролиферацию клеток, блокирует их клеточный цикл (Damiens et al., 1998). Индуцировать пролиферацию клеток в эксперименте можно, умеренно повышая тем или иным способом образование перекисей липидов, например, скармливанием подопытным животным в избытке того же железа (Whittaker et al., 1992).
Врeменные процессы активации и пролиферации клетки для своей реализации требуют, по-видимому, циклического возникновения умеренной гипероксии, повышенного на этот период пероксидативного напряжения и соответственно возрастания дисбаланса между ПО и АО частями его до значений, укладывающихся в диапазон DП (ПО – АО) и названных нами условно «пролиферативными». Данное представление в принципе отражает суть используемого в литературе термина «окислительный митогенез». Фактически это подтверждают и материалы обзора, авторы которого (Меньшикова, Зенков, 1997) после анализа многих публикаций последних лет пришли к заключению: «окислительный стресс средней интенсивности приводит к индукции клеточной пролиферации». Кроме того, представляется весьма вероятным, что такой стресс вообще распространён в биологическом мире и характерен в норме для совершенно далёких эволюционно видов. Например, показано: у спор N. crassa вскоре после поглощения ими О2, т.е. в начале их прорастания, развивается даже гипероксидантное состояние. На это указывало образование АФК, установленное путём анализа зависимой от О2 хемилюминесценции, окисления белков и некоторых других проявлений (Lledias et al., 1998).
Особенно наглядно временный характер окислительного митогенеза проявляется при регенерационных процессах. Так, после частичной гепатэктомии в процессе печёночной регенерации увеличиваются уровни липопероксидазы и ПОЛ, а содержание арахидоновой и других полиненасыщенных жирных кислот уменьшается. Для подавления ПОЛ в ранней фазе регенерации может быть использован витамин А. ПОЛ, таким образом, признаётся одним из сущест-венных факторов инициации регенерации печени (Kishino, 1998). В эмбрио-нальных же и стволовых клетках интенсивная пролиферация происходит вообще непрерывно. Ключевую роль в создании условий для такого окисли-тельного митогенеза играет, вероятнее всего, особенность митохондриальной базы указанных клеток: малое число митохондрий и относительно низкая сте-пень их дифференцированности. Впоследствии, по мере развития, содержание митохондрий увеличивается за каждый клеточный цикл. С тех же «мито-хондриальных» позиций обсуждаются и некоторые более общие вопросы взаимоотношения пролиферации и дифференцировки клетки (Von Wagenheim, Peterson, 1998).
В дальнейшем события могут развиваться по-разному в зависимости от складывающегося антиоксидантно-прооксидантного соотношения и его устойчивости в конкретной клетке. При некоторых превышающих DП (ПО – АО) значениях дисбаланса усиливаются процессы неферментативного образования АФК и обеспечивается более высокий уровень пероксидативного стресса. В случае длительного и непрерывного дестабилизирующего действия их на все компоненты клетки последняя может трансформироваться в злокачественную, т.е. в бесконтрольно размножающуюся с проявлением характерных для опухолевых клеток свойств. Происходящие при этом изменения в клетке должны быть сходны и в то же время более выражены по сравнению с теми, что имеют место при ее старении. Поэтому приуроченность онкологических заболеваний в большинстве случаев к позднему онтогенезу (Анисимов, 1986) неслучайна и может быть научно объяснена. Значения DК (ПО – АО), при которых нормальная клетка перерождается в опухолевую, будем считать «канцерогенными». К такому исходу в различных тканях организма приходит, по-видимому, лишь небольшой процент клеток, а установлению подходящих для трансформации условий способствует заметное к этому времени истощение в них эндогенной антиоксидантной системы и соответствующая «корректировка» комплекса метаболических и регуляторных процессов (см. главу 2). Для большинства же клеток и тканей возрастные изменения их направлены в сторону снижения пролиферативных потенций, затухания жизненного процесса. При этом возможны различные варианты регрессивного развития.
В одних клетках относительно быстрое возрастание значений параметра D (ПО – АО) приводит к превышению некоторого предельно допустимого уровня пероксидации и достижению значений DЦ (ПО – АО), ответственных за необратимую окислительную деградацию клетки, ее цитолиз. При этом варианте в постулируемой достаточно условной градации дисбалансы связаны между собой неравенствами
DИ (ПО – АО) < DП (ПО – АО) < DК (ПО – АО) < DЦ (ПО – АО).
Однако в действительности немалое количество клеток оказывается в состоянии гипоксии, вызванного известными при старении нарушениями в сосудистой системе и внешнем дыхании организма (Нагорный и др., 1963; Фролькис, 1984). При дефиците О2 в клетках деградируют митохондрии в соответствии с принципом: нефункционирование живой структуры есть способ её саморазрушения (Муханкин, 1989). Вместе с тем сокращение общей «мощности» митохондрий в клетке можно рассматривать как адаптивную реакцию на гипоксию и аноксию. Если степень сокращения митохондриальной базы слишком велика и энергообеспеченность серьёзно нарушена, то такая клетка, скорее всего, погибнет. Это крайний случай. Для многих же клеток более вероятна ситуация, когда утрата части митохондрий ведёт к некоторому снижению потребления О2, причём даже при слабых поступлении и утилизации О2 в зависимости от их соотношения в клетке могут устанавливаться разные уровни рО2 и, следовательно, D (ПО – АО), от низких и вплоть до ведущих к избыточной пероксидации. Значит, эта категория клеток, хотя и с замедленной скоростью, также будет проходить через все указанные выше состояния.
Что касается параметра DС (ПО – АО), связанного со старением, то он проявляет своё действие в широком интервале значений
DБ (t-τ) (ПО – АО) < DС (ПО – АО) < 

Данное неравенство подтверждает уже упомянутое выше: старение клеток, тканей и организма в целом начинается через какой-то временной интервал τ после рождения и затем продолжается до наступления канцерогенеза или естественной их смерти. Индекс (t-τ) при DБ означает как раз, что по времени t начало старения сдвинуто от базового дисбаланса DБ (ПО – АО) на величину τ – период постепенного возрастания этого дисбаланса до определённого поро-гового значения, с превышения которого начинается фактически заметный процесс старения. В любом случае этот процесс связан с возникновением в клетках окислительного стресса и образованием в них реактивных соединений О2 (Lundberg et al., 2000).
Следует подчеркнуть, что каждый из рассмотренных дисбалансов представляет определённый диапазон изменений, а не одну какую-то величину. Кроме того, для каждой ткани и даже клетки все эти переменные параметры индивидуальны по величинам и диапазону своего проявления. Искусственная коррекция каждого из указанных дисбалансов путем усиления антиоксидантной их части будет, очевидно, снижать эффективность необходимых в норме метаболических процессов в нестимулированной клетке, а также её активацию и пролиферацию. Действительно, некоторые антиоксиданты способны ингибировать митогенез и могут считаться антимитогенами (Садовникова, 1989). На процессы же старения и канцерогенеза усиление АО-составляющей должно оказывать тормозящее действие. Напротив, любой фактор, искусственно повышающий ПО-составляющую и уровень свободнорадикальных реакций (гипероксия, облучение, тепловое воздействие, обогащение пищи легкоокисляемыми жирами и др.), должен ускорять старение и увеличивать онколо-гический риск. Подтверждений на этот счёт получено много (см. п. 1.2 и 1.8; Лю, Шайхутдинов, 1991).
Различные аспекты приведённых выше общих положений развиваются и обосновываются нами на протяжении данной и всех последующих глав с привлечением фактических материалов из экспериментальной и клинической геронтологии и онкологии, физико-химической и молекулярной биологии.
Еще по теме окислительная модификация биологических структур:
- В В Е Д Е Н И Е
- 1.1.1. Биологическая эволюция
- окислительная модификация биологических структур
- 1.1.3. Кислородно-перекисная концепция старения
- 1.1.4. Вообще, суть и название большинства известных теорий старения
- 1.3.1. Внутренняя мембрана митохондрий
- 1.3.2. Особого внимания заслуживает взаимосвязь процесса старения человека и разных видов животных с разнообразными делециями мтДНК.
- 1.7.1. Атеросклероз
- 1.7.2. Сахарный диабет типа 2
- 1.7.3. Болезнь Альцгеймера
- 2.1.4. Превышение рО2 в клетке сверх уровня нормальных вариаций
- 2.1.13. Утверждение Варбурга о повышенной потенциальной способности опухоли к гликолизу
- 3.6. Биоэнергетические аспекты механизма пролиферациии дифференцировки клеток
- Биофизическая и биохимическая характеристики хрусталика
- Мембранотропные соединения как индукторы автолиза
- Положения, выносимые на защиту.
- ВВЕДЕНИЕ
- Параметры оценки процессов окислительной модификации белков в клинико-биохимическом мониторинге эндотоксикоза.
- Комплексный подход к изучению параметров окислительной модификации белков и уровня молекул средней массы в клиникобиохимическом анализе эндогенной интоксикации.