Структурно-функциональные особенности эндотелия человека в условиях моделируемой гипогравитации
Первая и, пожалуй, единственная проблема, возникшая в ходе исследования, - необходимость дополнительной отработки адекватных контролен. Это связано с тем, что в ходе клиностатирования помимо самого эффекта измененной гравитации присутствует, на первый взгляд несущественный, эффект постоянного перемешивания среды культивирования, особенно заметный при длительных экспериментах.
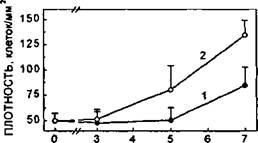
В статической культуральной системе, обычно используемой при работе с прикрепленными к подложке клетками, культуры достаточно быстро "кондиционируют" тонкий слой среды. Это связано, во-первых, с утилизацией содержащихся в ней питательных веществ и, во-вторых, с секрецией в среду продуктов клеточного метаболизма, в том числе биологически активных (цитокинов, ростовых факторов и т.п.). При этом даже при использовании избыточных (в пересчете на число клеток) объемов среды в статических условиях культуры реально используется лишь ее незначительная "донная" часть. Поэтому, для преодоления данного артефакта перемешивание среды культивирования в ходе длительных экспериментов становится необходимым. В качестве примера, демонстрирующего, насколько могут различаться функциональные параметры клеток в зависимости от условий культивирования, на рисунке 64 приведены кривые роста двух параллельных культур эндотелия. Кривая 1 соответствует клеткам, растущим в классической статической культуре; кривая 2 - тем же клеткам, но при регулярном перемешивании среды культивирования.
ВРЕМЯ КУЛЬТИВИРОВАНИЯ, сутки
Рисунок 64.
Сравнение пролиферативной активности ЭК человека, растущих в "стандартных" статических условиях (кривая 1) и при регулярном перемешивании среды культивирования (кривая 2).
Условия культивирования контрольных клеток, удовлетворяющие указанным требованиям, описаны в разделе "Материалы и методы".
8.1.Особенности пролиферации и формирования монослоя ЭК в условиях моделируемой гипогравитации
Одним из наиболее изученных проявлений ответа различных клеточных систем на измененное микроокружение является изменение скорости пролиферации клеток.
В совокупности с данными, полученными при исследовании эффектов гипо- и микрогравитации на пролиферативную активность других типов культивируемых клеток [Cogoli, 1997; Nash et al, 1991], это позволяет предположить, что если ЭК чувствительны к гравитационному стимулу, - они должны отвечать соответствующим изменением скорости роста. Для проверки данного предположения были исследованы некоторые параметры пролиферации и формирования монослоя в культуре ЭК в условиях длительного клиностатирования. Результаты одного из серии репрезентативных экспериментов представлены на рисунках 65 и 66.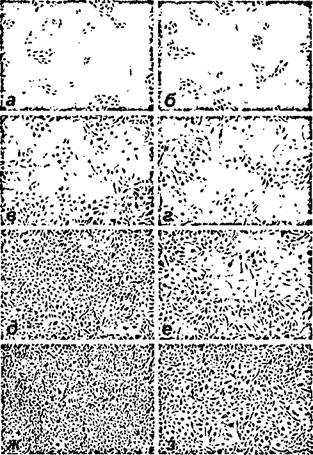
Рисунок 65.
Особенности роста и формирования монослоя ЭК человека при культивировании в контрольных условиях (а,в,д,ж) и при
клиностатировании (б.г.в.з). Микрофотографии соответствуют временным точкам: а, б - 2; в,г - 4; д,е - 6; ж,з - 8 суток.
Фазовый контраст, ув. 200Х.
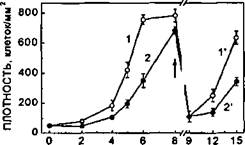
Полученные данные свидетельствуют, что скорость роста ЭК значительно (в среднем в 2 раза, различия статистически достоверны) снижена при воздействии гипогравитации по сравнению с контролем (рис. 66, кривые 1 и 2). Аналогичная
ВРЕМЯ КУЛЬТИВИРОВАНИЯ, сутки
Рисунок 66.
Пролиферативная активность ЭК человека в условиях гипогравитации in vitro. Кривые 1 и Г - контроль; кривые 2 и 2* - клетки культивировались в условиях клиностатирования при 5 об/мин. Стрелка указывает этап пересева клеток.
закономерность была выявлена и при пассировании культур, формирование монослоя в которых происходило в условиях предшествующей гипогравитации (Рис. 66, кривые Г и 2'. Стрелкой обозначен момент пересева культур). Как и в случае культур первого пассажа, стартовая плотность контрольных и опытных культур была идентична. Отчетливо видно, что сниженная пролиферативная активность ЭК в условиях гипогравитации сохраняется на протяжении всего времени субкультивирования.
Морфологический анализ культур позволил выявить еще одну интересную особенность формирования монослоя при клиностатировании. Клетки, растущие в условиях гипогравитации, выглядят более распластанными и достигают состояния прсконфлуэнтного монослоя (весь пластик уже покрыт клетками без промежутков между ними) при более низкой плотности, чем контрольные. Однако в последующие несколько суток плотность таких культур продолжает нарастать, и они практически догоняют контрольные. Максимальная плотность эндотелиальных монослоев, сформированных в контрольных культурах и культурах в условиях клиностатирования, по результатам нескольких экспериментов достоверно не отличается и составляет 750-800 клеток на 1 мм2. Возможно, сниженные темпы роста ЭК в условиях гипогравитации несколько компенсируются увеличением степени распластанности клеток, что способствует более раннему формированию межклеточных контактов, необходимых для нормального функционирования эндотелия.
Таким образом, данное исследование показало, что пролиферативная активность эндотелия человека значительно снижена в условиях длительного клиностатирования по сравнению с традиционными условиями культивирования.
Полученные результаты однозначно свидетельствуют, что, наряду с чувствительностью к механическим факторам [Shirinsky et al, 1989; Romanov et al, 1995], эндотелий человека способен к восприятию гравитационного микроокружения.
8.2.Влияние моделируемой гипогравитации на цитоскелет и миграционную активность клеток
В контрольных конфлуэнтных культурах ЭК, окрашенных на 5-7 сутки после посадки, F-актин был представлен организованной сетью филаментов, расположенных преимущественно в периферической части цитоплазмы и направленных вдоль границ клеток (Рис. 67 А). Часть клеток содержала умеренное количество волокон напряжения и звездчатых структур, характерных для клеток, культивируемых в присутствии дополнительных факторов роста [Smirnov et al, 1989]. Уже через 1-2 часа после начала клиностатирования актиновый цитоскелет претерпевал ряд заметных изменений: филаменты истончались и начинали перераспределяться в область межклеточных контактов (Рис.
67 Б). На протяжении 6-12 часов центральная часть клеток практически освобождалась от актиновых фибрилл и волокон напряжения. В то же время в краевой зоне наблюдалось формирование непрерывной линии F-актина, четко контрастирующей границы клеток (Рис. 67 В и Г). В последующие 12-24 часа существенных изменений не происходит.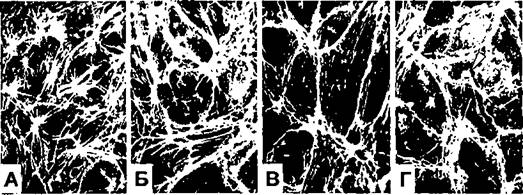
Рисунок 67. Реорганизация актиновых филаментов в культивируемых ЭК человека в условиях гипогравитации. А - контроль; Б - Г - клиностатирование в течение 4,12 и 18 часов, соответственно. Окрашивание РИТЦ- фаллоидином; Х450.
В большинстве ЭК описанные изменения цитоскелета сопровождались формированием так называемого "волнистого края" (ruffled edge), обогащенного F- актином (Рис. 67 Г; указано стрелкой). Поскольку и утрата волокон напряжения и формирование волнистого края способствуют усилению клеточной подвижности [Herman et al, 1981], миграционная активность клеток была исследована на хорошо зарекомендовавшей себя ранее модели репарации механически поврежденного монослоя ЭК [Smirnov et al, 1988]. Для этого, конфлузнтные монослои ЭК повреждали, нанося оплавленным концом стеклянного капилляра прямую царапину шириной от 200-300 до 500-600 мкм (примерно от 5-7 до 10-14 рядов клеток, соответственно). Из опыта предшествующей работы и литературных источников известно, что репарация подобных, незначительных по ширине участков повреждения эндотелия in vitro и in vivo, осуществляется в течение нескольких часов исключительно за счет распластывания и миграции клеток и не сопровождается всплеском пролиферативной активности [Hirsh, Robertson, 1977; Reidy, Schwartz, 1981; Smirnov et al, 1988; Романов, 1989]. Стадии реэндотелиализации поврежденного участка контролировали с помощью фазово-контрастной компьютеризированной видео-микроскопии. Временные интервалы, в которые проводилось фотографирование, указаны в подписях к соответствующим рисункам.
Вид культур до и после нанесения повреждения представлен на рисунке 68 А и Б, соответственно.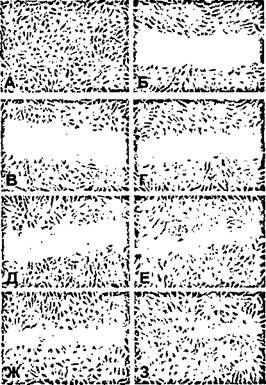
Рисунок 68.
Изменение скорости репарации механически поврежденного монослоя ЭК в условиях гипогравитации.
А, Б - интактный монослой ЭК и культура через 5 минут после нанесения повреждения, соответственно;
В - 3 - стадии репарации монослоя в контрольных условиях (слева) и при клиностатировании (справа).
Представленные фотографии соответствуют временным интервалам 30 минут (В, Г), 1 час (Д, Е) и 3 часа (Ж, 3).
Фазовый контраст, Х200.
Результаты одного из серии проведенных экспериментов представлены на рисунках 68 и 69. Видно, что уже в течение первых 30 минут в опытных культурах происходит активная миграция ЭК в область повреждения, сопровождающаяся репарацией до 50% деэндотелиализованного участка (Рис. 68 Г). В эти же сроки в контроле наблюдается лишь незначительная миграция клеток (Рис. 68 В). Различия в миграционной активности культур сохраняются и в последующие 2-3 часа (Рис. 68 Д-3). За это время в опытных культурах репарация области повреждения практически завершается: два фронта клеток, мигрирующих навстречу друг другу, смыкаются (Рис. 68 Е и 3). Напротив, в контрольных культурах область, не покрытая клетками, сохраняется на протяжении 3-4 часов (Рис. 68 Д и Ж), и лишь к концу 5-го часа зона повреждения закрывается полностью. В данном эксперименте опытные культуры были подвергнуты предварительному клиностатированию на протяжении 24 часов.
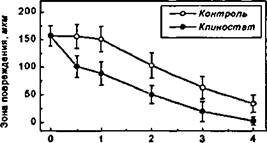
Время, ч
Рисунок 69.
Динамика репарации механически поврежденного монослоя ЭК в контрольных условиях и при клиностатировании.
Аналогичные результаты (2-3-кратное увеличение скорости миграции ЭК) были получены в экспериментах, где это время составляло 12 и 48 часов, а ширина области повреждения колебалась от 200 до 700 мкм.
В последнем случае различия между опытными и контрольными культурами сохранялись, соответственно варьировало лишь время, необходимое для репарации монослоя.Сходные с представленными изменения организации актинового цитоскелета ЭК, степени распластанности клеток и скорости репарации механически поврежденного монослоя были описаны нами ранее в ответ на добавление цАМФ- повышающих препаратов [Smirnov et al, 1988, 1989; Antonov et al, 1986]. Поэтому, полученные результаты позволяют предположить, что воздействие гипогравитации на эндотелий человека связано с изменением активности систем вторичных посредников, в частности, аденилатциклазы и сопровождается изменением многих звеньев клеточного метаболизма.
8.3.Экспрессия молекул клеточной адгезии и взаимодействие ЭК-лейкоцит в условиях гипогравитации
К моменту анализа адгезивных параметров клеток плотность контрольных и опытных культур составляла, соответственно, 814+61 и 805+54 клеток/мм2, т.е. достоверно не отличалась. Добавление TNF-a приводило к некоторой реорганизации монослоя в результате изменения формы клеток на более вытянутую, однако, изменения плотности культур при этом не наблюдалось.
За редким исключением, контрольные ЭК, выращенные в статических условиях, не содержали на своей поверхности определяемых количеств Е-селектина и VCAM-1 (Рис. 70 А, Б). Напротив, большинство клеток было положительно
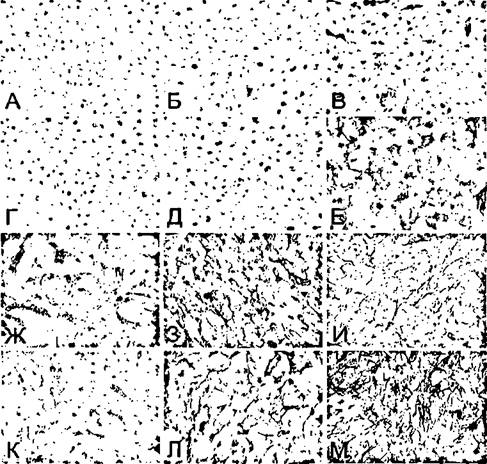
Рисунок 70. Влияние моделируемой гипогравитации на экспрессию Е-селектина (слева), VCAM-1 (в середине) и ICAM-1 (справа) в культуре эндотелиальных клеток человека.
А-В - статический контроль; Г-Е - клиностатирование в течение 18 ч;
Ж-И - активация ЭК с помощью TNF-a (10 ед./мл, 18 ч);
К-М - то же, в условиях клиностатирования. Иммунопероксидазная техника. Х200.
окрашено на ICAM-1, что отчетливо видно на препаратах, окрашенных с использованием пероксидазной техники (Рис. 70 В). Добавление цитокинов, интерлейкина-1 или TNF-a (5-10 ед/мл) уже в течение 6-8 часов индуцировало экспрессию Е-селектина и VCAM-1 и приводило к значительному повышению клеточного содержания ICAM-1 (Рис. 70 Ж-И). Поскольку выявленная активность цитокинов не отличалась от описанной в предшествующих разделах работы, временные зависимости не приводятся.
Через 24 часа от начала клиностатирования клетки были по-прежнему негативны в отношении экспрессии Е-селектина и VCAM-1. В то же время интенсивность окрашивания на ICAM-1 значительно возрастала (Рис. 70 Г-Е). Наиболее яркое окрашивание на этот белок было выявлено при клиностатировании клеток в присутствии TNF-a (Рис. 70 М).
Если клиностатирование способствовало спонтанной и потенцировало индуцированную цитокинами экспрессию ICAM-1, то в отношении Е-селектина и VCAM-1 картина была противоположной. В ответ на добавление цитокинов и интенсивность окрашивания, и число положительных клеток (при анализе с помощью поточной цитофлуориметрии) были значительно ниже при клиностатировании, нежели на активацию теми же концентрациями агентов в
в
статических условиях (Рис. 70 Ж,3,К,Л). Результаты проточной цитофлуориметрия представлены в таблице 8.
| контроль | клиностат | TNF-a | TNF-a +клиностат | |
| Е-селектин | 4.0 | 3.1 | 55.6 | 32.3 |
| VCAM-1 | 3.4 | 2.4 | 78.4 | 52.7 |
| ICAM-1 | 30.9 | 48.6 | 82.9 | 97.7 |
Таблица 8. Изменение числа (%) положительно окрашенных клеток при клиностатировании клеток в присутствии или отсутствии дополнительной активации.
Результаты анализа изменений адгезивных параметров ЭК в контрольных, активированных цитокинами и клиностатирусмых культурах приведены на рисунках 71 и 72. Лимфоциты слабо взаимодействовали с контрольными клетками и практически не адгезировали на ЭК, подвергнутые клиностатированию (8.5+2.0 и 6.1+2.1 лейкоцитов на 100 ЭК, соответственно, p=NS). Активация клеток крови с помощью ФМА заметно повышала их способность взаимодействовать с эндотелием, особенно, подвергнутым клиностатированию (1.19+0.13 и 1.45+0.13 лимфоцитов на 1 ЭК, соответственно, р
Еще по теме Структурно-функциональные особенности эндотелия человека в условиях моделируемой гипогравитации:
- 1. Особенности организации эндотелия аорты человека в норме и при атеросклерозе
- Структурные и функциональные особенности генов В диагностике и лечении рака
- Морфологическая гетерогенность эндотелия человека
- Поиск функциональных различий эндотелия в зонах с различной
- Метод оценки функционального состояния и функционального резерва организма и его систем в условиях действия электромагнитных факторов.
- 4.2. Функциональная гетерогенность популяций эндотелия
- Изучение организации эндотелия аорты человека in situ
- 4.1 Функционально-структурная организация ИАСППР
- 1.2.3. Влияние оксидативного стресса и воспаления на функциональную активность эндотелия
- Структурные составные части и основные функциональные процессы
- Структурные и функциональные предпосылки сознания