Экспрессия МКА и взаимодействие эндотелий-лейкоцит в культуре
Согласно современным представлениям, молекулы клеточной адгезии (МКА) являются одним из ключевых звеньев в регуляции межклеточных взаимодействий: и морфогенез в эмбриональном периоде, и заживление повреждения тканей, и воспаление непосредственно связаны с экспрессией этих белков [Ginsberg et al, 1988; Gimbrone et al, 1990; Takeichi, 1991; Edelman, 1992].
Такие процессы, как метастазирование опухолей и атерогенез, также не обходятся без участия МКА [Cybulsky, Gimbrone, 1991; Taichman et al, 1991]. Во взаимодействии клеток крови, в частности, с сосудистым эндотелием принимают участие, по крайней мере, три основных класса МКА [Bevilacqua, 1993]. К первому, семейству селсктинов, относятся Р-селекгин (GMP-140) и Е-селектин (ELAM-1), определяющие кратковременное взаимодействие клетки крови с эндотелиальной клеткой и так называемый "роллинг" по луминальной поверхности сосудов [Hogg, 1992; Patel et al, 1993]. Ко второму, суперсемейству иммуноглобулинов, относятся ICAM-1, VCAM-1, ICAM-2 и РЕСАМ-1, участвующие в формировании более прочного межклеточного контакта и миграции лейкоцитов в субэндотелиальное пространство [Bevilacqua, 1993; Muller et al, 1993]. И, наконец, третью группу составляют интегрины на поверхности клетки крови, являющиеся специфическими рецепторами для соответствующих МКА на мембране ЭК [Hynes, 1992; Bevilacqua, 1993]. Характерной особенностью неактивированного эндотелия является отсутствие (за исключением ICAM-1) МКА на поверхности клеток. Р-селектин, хотя и содержится в тельцах Вейбеля-Паладе, лишь кратковременно экспрессируется на клеточной поверхности при активации ЭК тромбином или гистамином и затем вновьинтернализуется [Patel, et al, 1993]. Е-селектин и VCAM-1 не имеют внутриклеточных хранилищ и синтезируются de novo только в ответ на активацию цитокинами [Bevilacqua, 1993]. Содержание ICAM-1 в этих условиях также возрастает.
В результате, в нормальных условиях in vivo и in vitro эндотелиальный пласт остается атромбогенным и неадгезивным для клеток периферической крови. Напротив, в участках микровоспатения и на ранних стадиях атерогенеза ситуация, по-видимому, кардинально изменяется.Немногочисленные исследования экспрессии различных классов МКА эндотелием, выстилающим области атеросклеротических поражений сосудов животных и человека, показали присутствие в эндотелиальной выстилке участков, положительно окрашенных на эти белки [Cybulsky, Gimbrone, 1991; Poston et al, 1992; O’Brien et al, 1993; Wood et al, 1993; Johnson-Tidey et al, 1994; Walpola et al, 1995]. Одновременно, в субэндотелии обнаруживались скопления клеток гематогенного происхождения (макрофагов, лимфоцитов и т.д.). Тем не менее, авторы этих работ не смогли однозначно ответить на вопрос, какое же событие является первичным: либо повышенная экспрессия МКА в этих участках определяет последующую инфильтрацию клетками крови, либо присутствие последних и секретируемые ими цитокины активируют вышележащие ЭК.
С другой стороны, даже в отсутствие инфильтрации субэндотелия клетками крови, интимальные ГМК могут, по-видимому, также рассматриваться, как один из источников цитокинов в сосудистой стенке, поскольку способны секретировать значительные количества интерлейкина-1 (ИЛ-1) и фактора некроза опухолей-альфа (TNF-a), являющихся регуляторами экспрессии МКА эндотелием [Thornhill et al, 1991; Bevilacqua, 1993]. С этой позиции именно ГМК могут являться клеточным элементом, способным (в отсутствие других участников) влиять на экспрессию МКА ЭК и их последующее взаимодействие с клетками периферической крови.
Для проверки данного предположения была исследована возможность участия интимальных ГМК в регуляции экспрессии Е-селектина, ICAM-1 и VCAM-1 в культуре ЭК человека. Контрольные, т.е. не активированные цитокинами ЭК, не содержали на своей поверхности ни Е-селектина, ни VCAM-1 (Рис. 36 А, В). Напротив, практически все клетки несли на своей поверхности незначительное количество 1САМ-1 (Рис.
36 Б). Добавление ИЛ-1 или TNF-a в концентрации 2-20 ед./мл приводило к появлению на клеточной мембране Е-селектина и VCAM-1 и значительному усилению экспрессии ICAM-1 (Рис. 37). Как при иммунопероксидазном окрашивании, так и при поточной цитофлуориметрии было установлено, что эффекты цитокинов проявляются уже через 2-4 ч и достигаютмаксимума через 6-8 ч от начала воздействия. В последующие 12-24 ч концентрация Е-селектина снижалась до базального уровня, тогда как VCAM-1 и ICAM-1 сохранялись на клеточной поверхности в высокой концентрации в течение всего времени присутствия активатора в среде культивирования, что согласуется с известными литературными данными.
ИНТЕНСИВНОСТЬ ФЛУОРЕСЦЕНЦИИ
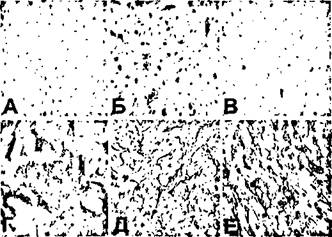
Рисунок 36.
Изменение экспрессии Е-селектина (А, Г). 1САМ-1(Б,Д) и VCAM-1 (В,Е) при со-культивировании ЭК и ГМК в системе TransWell.
А-В - контроль;
Г-Е - со-культивирование. Авидин-биотиновая техника. Х400.
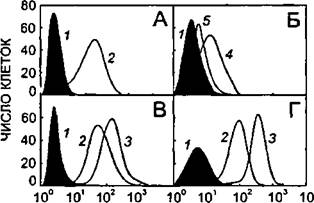
Рисунок 37.
Изменение содержания Е- селектина (А, Б), VCAM-1 (В) и ICAM-1 (Г) на поверхности ЭК, инкубированных в течение 8 часов в присутствии различных активаторов.
1 - контрольные клетки;
2 - со-культивирование с ГМК;
3 - КС-ГМК;
4- ИЛ-1, 2.5 ед./мл;
5 - КС-ГМК + ИЛ-1, 2.5 ед./мл.
Добавление к культивируемым ЭК кондиционированной среды от ГМК (КС- ГМК), равно как помещение в ячейку с ЭК вкладыша Transwell с ГМК, приводило к выраженным изменениям содержания МКА на клеточной поверхности (Рис. 37; Рис. 38). В случае VCAM-1 и ICAM-1 эффект со-культивирования был аналогичен воздействию на клетки ИЛ-1, взятого в концентрации 5-10 ед. на 1 мл среды. Как и в первом случае, изменения становились заметными через 2-4 ч и достигали максимума через 8-12 ч (Рис. 38). Отличием было то, что при со-культивировании нарастание концентрации МКА происходило медленнее, чем при добавлении кондиционированной среды.
Кроме этого, при добавлении КС-ГМК экспрессия
VCAM-1 возрастала и достигала максимума через 8-12 ч, а затем снижалась, чего не происходило при добавлении ИЛ-1 или при со-культивировании двух типов клеток.
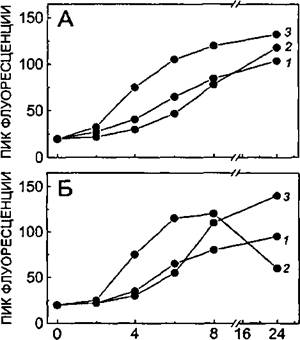
Рисунок 38.
Динамика изменения содержания ICAM-1 (А) и VCAM-1 (Б) при активации ЭК различными индукторами:
1 - ИЛ-1 (5 ед./мл);
2- КС-ГМК;
3- со-культивирование с ГМК.
В отличие от VCAM-1 и ICAM-1, изменения экспрессии Е-селектина под воздействием КС-ГМК и в результате со-культивирования оказались различными по направленности действия. Так, при со-культивировании Е-селектин начинал выявляться на клеточной поверхности через 2-3 ч, в последующие 4-12 ч его содержание нарастало, достигало максимума и сохранялось на высоком уровне вплоть до 24 часов (Рис. 37 А). Добавление кондиционированной среды, напротив, не только не индуцировало появление этого белка, но и практически полностью блокировало экспрессию, вызванную добавлением низких доз ИЛ-1 (0.5-2.5 ед/мл), а также в значительной мере снижало эффект более высоких (5-10 ед/мл) концентраций цитокина (Рис. 37 Б).
Тем не менее, и добавление КС-ГМК и со-культивирование двух типов клеток резко повышали адгезивные параметры ЭК по отношению к линии HL-60. Так, если адгезия клеток HL-60 на контрольные ЭК была минимальна (10-15 клеток на 1 см2 монослоя) и лишь незначительно повышалась при активации промиелоцитов хемотаксическим пептидом FMLP или форболовым эфиром ФМА, то в случае использования кондиционированной среды этот показатель возрастал до 250-300 клеток на 1 мм2 (Рис. 39).
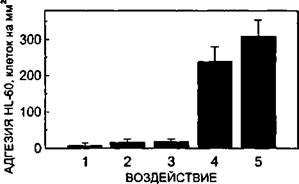
Рисунок 39.
Адгезия клеток линии HL-60 на ЭК при активации эндотелия КС-ГМК. 1 - контроль; 2, 3-то же, но клетки HL-60 были активированы FMLP (30 нМ) и ФМА (100 нМ), соответственно; 4 - после активации ЭК КС-ГМК в течение 18 ч; 5 - то же, после активации клеток HL-60 пептидом FMLP.
Таким образом, в ходе проведенных экспериментов удалось показать, что, по крайней мере, в условиях культивирования in vitro ГМК способны кардинально изменить экспрессию МКА эндотелием. В отношении VCAM-1 и ICAM-1 биологическое действие кондиционированной среды от ГМК или их эффект в условиях со-культивирования были приблизительно одинаковы и сравнимы по силе с непосредственным добавлением в среду культивирования достаточно высоких (5- 10 ед./мл) концентраций ИЛ-1. Одновременно с усилением экспрессии этих белков на поверхности ЭК резко возрастала и адгезия клеток HL-60 на эндотелий.
Кроме этого, выяснилось, что, в зависимости от характера воздействия, цитокины, секретируемые ГМК, могут, как инициировать, так и блокировать экспрессию Е-селектина на поверхности эндотелия. При этом если активаторами синтеза этого белка могут служить те же ИЛ-1 и TNF-a, то наиболее вероятным агентом, блокирующим эту экспрессию, является трансформирующий ростовой фактор-бета (TGF-P), синтезируемый ГМК в культуре и in situ. Основанием для этого являются экспериментальные данные, показавшие, что TGF-P способен блокировать экспрессию Е-селектина в активированных цитокинами ЭК и снижать их адгезивные свойства [Gamble, Vadas, 1988, 1991; Gamble etal, 1993].
Таким образом, полученные экспериментальные данные свидетельствуют, что интимальные ГМК являются мощным регулятором эндотелиальных функций, причем сила и направленность воздействия на эндотелий может зависеть не только от их секреторной активности в целом, но и от баланса секретируемых цитокинов.
6.
Еще по теме Экспрессия МКА и взаимодействие эндотелий-лейкоцит в культуре:
- Другие участники взаимодействия ЭК - лейкоцит
- Содержание многоядерных ЭК в культурах эндотелия из зон НПА и ВПА
- Амплификация полной кодирующей последовательности целевого гена. Клонирование и экспрессия гена флуоресцентного белка в бактериальной системе экспрессии (задача 4)
- Фактор роста эндотелия сосудов
- Морфологическая гетерогенность эндотелия человека
- Повреждение и репарация эндотелия
- Пространственная организация эндотелия в зонах НПА и ВПА.
- Поиск функциональных различий эндотелия в зонах с различной
- Эндотелий атеросклеротической аорты
- Частота выявления кластеризованного эндотелия в зонах НПА и ВПА
- Выбор системы экспрессии
- Экспресс-тесты на ВИЧ.
- 1. Особенности организации эндотелия аорты человека в норме и при атеросклерозе
- Возрастная динамика изменений эндотелия визуально неизмененных сосудов