Молекулярно-генетическая модель таупатии на основе сверхэкспрессии протеинкиназы GSK3p в нервной системе Drosophila melanogaster
М.В. Тростников, Н.В. Рощина, А.В. Симоненко, Д.В. Муха, Е.Г Пасюкова
Снижение смертности и продление здоровой человеческой жизни являются одним из приоритетных направлений работы в биологии и медицине.
Известно, что старение сопряжено с развитием различных нейродегенеративных заболеваний, и следовательно предупреждение и лечение этих заболеваний, замедление старения и продление жизни могут быть связаны с коррекцией одних и тех же метаболических путей и вызваны вмешательством одних и тех же агентов.Болезнь Альцгеймера является одним из наиболее распостраненных нейродегенеративных заболеваний, сопровождающихся деменцией, причем одним из основых ее призаков является гиперфосфорилирование и накопление белка тау в нейронах и клетках глии (Buerger et al., 2006; Ramachandran, Udgaonkar, 2013; Mal’tsev et al., 2013; Pooler et al., 2013). Таупатия возникает также при развитии другого нейродегенеративного заболевания - болезни Паркинсона, которая также сопровождается деменцией (Kawakami et al., 2013; Compta et al., 2013). Этот же диагностический признак характерен и для других, более редких болезней, сопровождающихся развитием слабоумия - болезни Пика, лобно-височной деменции, прогрессирующего надъядерного паралича, кортико-базальной дегенерации и некоторых других (Rowe et al., 2007; Noble et al., 2013; Pan, Chen, 2013). В настоящее время считается, что именно уровень фосфорилирования тау определяет развитие таупатий (Noble et al., 2013). Фосфорилирование белка тау обусловлено действием нескольких протеинкиназ, среди которых ключевую роль играет серин-треонино- вая протеинкиназа GSK-3 P (glycogensynthase kinase-3 Р) (Noble et al., 2013). В связи с этим ингибирование GSK-3 P можно рассматривать как потенциальное средство борьбы с таупатиями (Rockenstein et al., 2007). Так, ингибирование GSK-3 P литием уже используется для лечения маниакального синдрома и некоторых других болезней (Rowe et al., 2007; Medina, Avila, 2014).
Протенкиназа GSK-3 P представляет собой мультифункциональный высококонсервативный белок, участвующий в регуляции сходных процессов у различных организмов, включая дрозофилу и человека. Взаимодействуя с Wnt-каскадом (Ataman et al., 2008; Chiang et al., 2009), GSK-3 P играет важную роль в развитии нервной системы, определяет активность синапсов, поведение, память. GSK-3 P связана также с функционированием инсулинового и TOR (Target of Rapamycin) каскадов - ключевых путей регуляции гомеостаза, метаболизма и старения (Kapahi et al., 2010). Интересно, что активация инсулинового и Wnt-каскадов снижает уровень фосфорилирования тау, опосредованный GSK-3 P (Medina, Avila, 2014). Избыточная акивность GSK-3 P в нервной системе приводит не только к гиперфосфорилированию тау, но также влияет на процессинг APP (Amyloid Precursor Protein ) и образование амилоидных пептидов, регулируя транскрипцию BACE1 (P-Secretase 1), изменяет свойства ионных каналов, вызывает нарушение функций митохондрий (Medina, Avila, 2014; Reddy, 2013). Существенную роль играет GSK-3 P в развитии различных других заболеваний - воспалений, диабета, ожирения (Reddy, 2013).
В последние годы для исследования молекулярно-генетических основ различных патологий и скрининга потенциальных лекарств всё большее использование находят модели болезней человека, созданные с использованием Drosophila melanogaster (Pandey, Nichols, 2011; Konsolaki, 2013). Высокий потенциал этих моделей обусловлен их высокой специфичностью, а также особенностями Drosophila melanogaster как объекта, пригодного для проведения умеренно массовых скринингов потенциальных лекарств (Rockenstein et al., 2007). Как модельный организм Drosophila melanogaster имеет множество преимуществ. В ее геноме, меньшем, но в то же время сравнимом по размеру с геномом человека, около 70% генов являются ортологами генов человека, а многие клеточные и физиологические процессы сходны на молекулярном уровне. Для Drosophila melanogaster разработан наиболее изощренный по сравнению с другими модельными организмами генетический инструментарий.
В распоряжении исследователей находятся мутации практически всех генов, работа около 80% генов может быть заблокирована с помощью РНК- интерференции, усилена или ослаблена в целой мухе и в отдельном органе или ткани благодаря использованию специальных коллекций линий (http://flybase.org/). Сравнительно короткие жизненный цикл и продолжительность жизни Drosophila melanogaster позволяют эффективно проводить эксперименты, требующие большого числа поколений, и многократно проводить опыты большого масштаба, необходимые для получения достоверных результатов; проведение таких опытов с млекопитающими имеет ряд технических и финансовых ограничений.В настоящее время у Drosophila melanogaster описано более 30 моделей различных болезней человека. Наибольший успех достигнут в создании моделей различных нейродегенеративных заболеваний (см. обзоры Cauchi, Van den Heuvel, 2006; Lu, Vogel, 2009; Jeibmann, Paulus, 2009), что обусловлено принципиальным сходством в развитии и структуре нейронов и системах передачи нервного импульса у Drosophila melanogaster и человека, а также возможностью адекватной оценки изменений в поведении и долгосрочной и краткосрочной памяти. Разработаны также модели различных онкологических заболеваний (Froldi et al., 2008; Palomero et al., 2008; Read et al., 2009), болезней сердечной мышцы (Wolf et al., 2006; Xu et al., 2006; Shcherbata et al., 2007; Taghli-Lamallem et al., 2008), инфекционных поражений (Nehme et al., 2007; Sibley et al., 2008; Dionne, Schneider, 2008), астмы (Roeder et al., 2009) и других болезней.
Таупатии были успешно смоделированы у Drosophila melanogaster с помощью сверхэкспрессии у мух гена белка тау быка, нормального и мутантного гена человека и гена Drosophila melanogaster (Williams et al., 2000; Wittmann et al., 2001; Jackson et al., 2002; Mershin et al., 2004). Эти модели хорошо воспроизводят основные черты таупатий человека: гиперфосфорилирование белка тау и его накопление, образование нейрофибриллярных клубков, нейродегенерацию, нарушение функции синапсов, сокращение продолжительности жизни.
Различные модели отличаются не только происхождением сверхэкспрессированного гена, но и тканью, где индуцировали сверхэкспрессию: весь организм, все клетки мозга, мотонейроны или другие отдельные группы нервных клеток и др. Эффект сверхэкспрессии гена тау усиливался, если одновременно в клетках сверхэкспрессировали ген белка GSK-3 P и уменьшался при использовании ингибиторов протеинкиназы (Mudher et al., 2004; Yeh et al., 2010). Сверэкспрессия генов тау в клетках ретины приводила к появлению фенотипа грубых глаз (Chen et al., 2007). Сверхэкспрессия гена белка тау человека в мотонейронах Drosophila melanogaster нарушала подвижность мух (Jackson et al., 2002; Mudher et al., 2004). Сверэкспрессия генов тау в дорзальной части мезоторакса приводила к потере щетинок (Yeh et al., 2010). Видно, что помимо нарушений структуры и функции мозга таупатия у Drosophila melanogaster характеризуется различными нарушениями фенотипа, которые могут служить диагностическими признаками для оценки токсичности тау в скринингах потенциальных лекарств.Создание лекарственных средств нового поколения, базирующееся на понимании механизмов болезни и научно обоснованных способов вмешательства в них, все шире распространяется за рубежом и начинает развиваться в нашей стране. Специфичность и направленность действия лекарства имеет принципиальное значение при лечении заболевания. Поиск терапевтических агентов и их мишеней, которые специфично взаимодействуют между собой, представляет собой сложную и актуальную задачу. Эффективное решение этой задачи становится возможным в результате разработки методов высокопродуктивного, чувствительного, быстрого и дешевого скрининга лекарственных препаратов. В основе такого скрининга лежит использование модельных систем различных заболеваний, позволяющих тестировать и отбирать препараты, обладающие заданной специфичностью в отношении конкретной патологии и даже конкретного механизма, вызывающего эту патологию. Использование модельных систем, позволяющих вести скрининг in vivo, является необходимым этапом поиска лекарств.
В связи с этим разработка таких модельных систем представляет собой важную и актуальную задачу.Преимущества поиска лекарственных средств с использованием Drosophila melanogaster основаны на следующем. 1) Возможность создания адекватных и гибких моделей болезней человека. 2) Возможность исследовать эффекты потенциальных лекарств на уровне целого организма и органов, свойства которых не удается воспроизвести у бактерий, дрожжей и на культуре клеток млекопитающих. 3) Простота и относительно малая стоимость содержания Drosophila melanogaster, короткий жизненный цикл. 4) Наличие технического оснащения для анализа диагностических признаков. Два последних пункта обусловливают возможность проведения умеренно массовых скринингов (Rockenstein et al., 2007). Первые попытки скринирования библиотек химических веществ с помощью моделей успешно осуществлены (Agrawal et al., 2005; Lai et al., 2008).
Целью нашей работы является создание молекулярно-генетической модели таупатии на основе трансгенных конструкций, обеспечивающих сверхэкспрессию различных форм протеинкиназы GSK-3 P в нервной системе Drosophila melanogaster. Эта модель может быть использована как инструмент для решения фундаментальных задач - исследования молекулярно-генетических основ функционирования нервной системы в норме и патологии и контроля продолжительности жизни. Она дополнит ряд уже существующих моделей и позволит детально исследовать роль GSK-3 P в этих процессах. Такая задача представляется чрезвычайно важной, поскольку стало очевидно, что эта роль сложна и многообразна (Sofola et al., 2010; Reddy, 2013; Medina, Avila, 2014), а взаимосвязь различных процессов, в которых участвует GSK-3 Р, и молекулярные механизмы их сопряжения остаются неясными.
Созданная модель таупатии также может быть адаптирована для практического использования в качестве тест-системы для скрининга потенциальных лекарственных препаратов. Использование такой модели позволит повысить специфичность скрининга и последующей терапии в отношении биомишеней, вызвающих гиперфосфорилирование тау.
Для осуществления поставленной цели в 2012-2014 гг. была заложена основа создаваемой модели: клонирована кДНК, соответствующая различным вариантам транскриптов гена shaggy, кодирующего GSK-3 Р, получены линии-трансформанты, необходимые для обеспечения сверхэкспрессии различных вариантов протеинкиназы GSK-3 P и гиперфосфорилирования белка тау в клетках мозга, предварительно охарактеризована экспрессия GSK-3 Р, подвижность, продолжительность жизни и функция синапсов у модельных особей.
В соответствии с принципиальной схемой модели гиперфосфорилирование тау достигается в результате увеличения уровня экспрессии GSK-3 P в нервной системе. Усиление экспрессии GSK-3 P достигается вследствие присутствия в геноме модельных мух дополнительной копии гена, кодирующего эту протеинкиназу, в составе специальной векторной конструкции. Экспрессия дополнительной копии гена (трансгена) обеспечивается благодаря использованию бинарной экспрессионной системы GAL4-UAS. Векторная конструкция помимо трансгена содержит дрожжевую последовательность UAS, активация которой с помощью специфического дрожжевого транскрипционного фактора GAL4 позволяет индуцировать экспрессию трансгена. Для активации экспрессии векторные конструкции (одна из них содержит дополнительную копию гена и UAS, а другая ДНК транскрипционного фактора GAL4 под управлением специфического промотора, обеспечивающего экспрессию в выбранной ткани) совмещаются в одном геноме в результате скрещивания.
Ген shaggy, кодирующий GSK-3 P дрозофилы, образует 17 различных транскриптов, которым соответствуют 11 форм белка (http://flybase.org). Для дальнейшего исследования было выбрано четыре транскрипта (рис. 1), существование которых было не только предсказано, но и продемонстрировано экспериментально (http:// flybase.org). Транскрипт RB (длина несплайсированной формы 2847 п.н., сплайси- рованной - 1545 п.н.) кодирует основную форму белка, PB. Функциональная роль этой формы белка описана в ряде работ (Bourouis, 2002; Ataman et al., 2008; Chiang et al., 2009). Транскрипт RA (длина несплайсированной формы 2929 п.н., сплайси- рованной - 1728 п.н.) кодирует форму белка PA. Транскрипт RG (длина несплайсированной формы 3174 п.н., сплайсированной - 1491 п.н.) кодирует форму белка
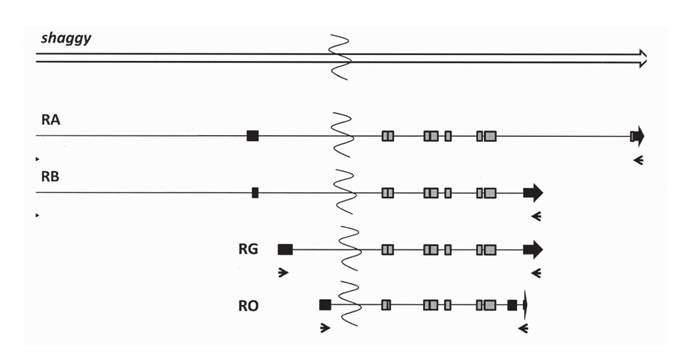
Рис. 1. Транскрипты гена shaggy
Интроны обозначены прямыми линиями, нетранслируемые области - черными прямоугольникам, транслируемые области - серыми прямоугольниками; стрелки обозначают позиции праймеров, использованных для получения к ДНК
PG. Транскрипт RO (длина несплайсированной формы 2166 п.н., сплайсированной - 1326 п.н.) кодирует форму белка PO. Функциональная роль этих форм белка неизвестна.
Анализ базы данных http://flybase.org показал, что линии, содержащие инсер- цию векторной конструкции, в состав которой входит кДНК, соответствующая нормальному варианту основной формы белка GSK-3 в, PB, а также вариантам с аминокислотными заменами в N-терминальном ингибирующем сайте, активационной петле и киназном домене (Bourouis, 2002), имеются в центре культур дрозофилы (Блюмингтон, США). Эти линии были выписаны, получены и две из них подготовлены для дальнейшей работы. Одна из линий, w(1118); P{w(+mC) = UAS-sgg.B}MB5, содержит инсерцию векторной конструкции, в состав которой входит кДНК, соответствующая нормальному белку PB, и последовательность индуцибельного дрожжевого промотора UAS (Bourouis, 2002). Вторая линия, w(1118); P{w(+mC) = UAS- sgg.Y214F}2, содержит инсерцию векторной конструкции, в состав которой входит кДНК, соответствующая белку PB с аминокислотной заменой Y214F в активационной петле, и последовательность дрожжевого промотора UAS. В результате мутации экспрессия трансгенной копии GSK-3 в снижена примерно в 20 раз (Bourouis, 2002). Таким образом, использование этих двух линий дает возможность сильного и умеренного индуцированного увеличения активности GSK-3 в.
Получение линий дрозофилы, содержащих инсерции векторных конструкций, в состав которых входит кДНК, соответствующая формам белка PA, PG и PO, было проведено нами. Тотальная РНК была выделена из целых мух, самцов и самок, линии, использованной ранее для полногеномного секвенирования (http://flybase. org). Геном именно этой линии аннотирован в базе данных, из которой были получены сведения о транскриптах и белках, кодируемых геном shaggy. Выделенная РНК была использована для получения кДНК. С помощью специфических праймеров (см. рис. 1) фрагменты кДНК, соответствующие сплайсированным формам RA, RG и RO, были амплифицированы и клонированы в плазмидном векторе pUC19. Было проведено секвенирование полученных вставок, с тем чтобы выявить клоны, в которых последовательность ДНК не имела возникших в ходе амплификации нуклеотидных замен по сравнению с референсной последовательностью из базы данных. Один такой клон из трех секвенированных был выявлен для RA, один из трех секвенированных - для RG, и один из шести секвенированных - для RO. Эти три клона были далее использованы для переклонирования в вектор для эмбриональной трансформации дрозофилы pBI-UASC.
Для переклонирования в вектор для эмбриональной трансформации дрозофилы pBI-UASC (http://www.columbia.edu/cu/mccabelab/PubHcations/Vectors) (рис. 2) вставки ДНК, соответствующие RA, RG и RO, были амплифицированы таким образом, что к их 5’ концу в положении, предшествующем стартовому кодону, была пришита последовательность Kozak CAAA, характерная для дрозофилы, для обеспечения лучшей экспрессии. Кроме того, к обоим концам были пришиты после-
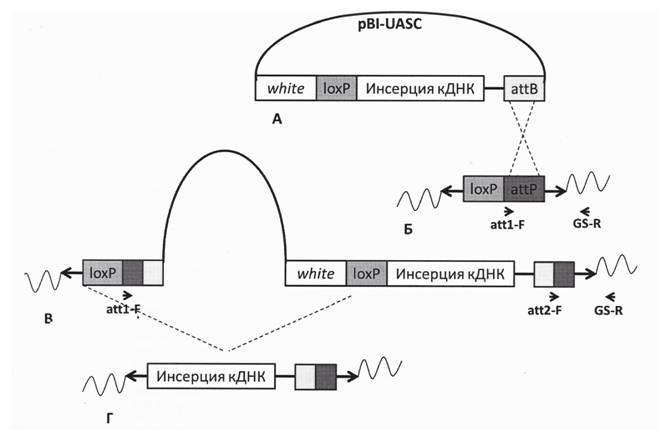
Рис. 2. Схема встраивания трансгенной конструкции в геном Drosophila melanogaster А - плазмида с кДНК копией гена shaggy. Б - посадочный сайт для интеграции в геном. В - полный вектор, интегрированный в геном. Г - вектор, интегрированный в геном, после вырезания вспомогательных последовательностей. white - ген-маркер эмбриональной трансформации; аЦВи attP - последовательности сайт-специфической интеграции фага PhiC31; IoxP - мишени для рекомбиназы фага P1; стрелки обозначают позиции праймеров (объяснения см. в тексте)
довательности, обеспечивающие клонирование в вектор pBI-UASC по выбранным сайтам рестрикции. Последовательность полученных в результате переклониро- вания вставок была проверена секвенированием. Полученные конструкции были далее использованы для получения линий-трансформантов.
Для эмбриональной трансформации дрозофилы была использована интеграционная система на основе фага PhiC31. Вектор pBI-UASC содержит attB-сайт, обеспечивающий сайт-специфическую интеграцию чужеродного генетического материала в районы хромосом дрозофилы с заранее интегрированым attР-сайтом (см. рис. 2). Интеграза фага осуществляет рекомбинацию между att-сайтами и обеспечивает встройку. Для трансформации была выбрана линия y(1) M{vas-int. Dm}ZH-2A w(*); M{3xP3-RFP.attP'}ZH-51C, которая содержит встроенную векторную конструкцию, кодирующую интегразу, встроенную векторную конструкцию с attP-сайтом, и характеризуется высокой эффективностью трансформации и отсутствием влияния геномных последовательностей на уровень экспрессии трансгена. Для каждой формы белка GSK-3 P (PA, PG и PO) было отобрано по несколько мух-трансформантов, из которых были заложены линии. Правильность встройки конструкции в каждой линии была подтверждена с помощью амплификации фрагмента ДНК с праймеров, один из которых гомологичен геномному окружению (GSR), а второй - встроенной последовательности (att1-F, см. рис. 2). Далее, индукция рекомбинации по встроенным в трансгенные конструкции loxP-сайтам позволила вырезать из генома большую часть вспомогательных последовательностей, оставив в составе хромосом только трансгены (см. рис. 2). Эффективность вырезания была проверена с помощью ПЦР с праймерами att1-F, att2-F и GS-R (см. рис. 2). Для дальнейшей работы были выбраны три линии: y(1) M{vas-int.Dm}ZH-2A w(*); M{3xP3-RFP.attP'}ZH-51C P{w(+mC)=UAS-sgg.A}1M - содержит трансген, соответствующий форме PA; y(1) M{vas-int.Dm}ZH-2A w(*); M{3xP3-RFP.attP'}ZH-51C P{w(+mC)=UAS-sgg.G}1M- содержит трансген, соответствующий форме PG, y(1) M{vas-int.Dm}ZH-2A w(*); M{3xP3-RFP.attP'}ZH-51C P{w(+mC)=UAS-sgg.O}1M - содержит трансген, соответствующий форме PO.
Для создания модели был также сформирован набор специальных линий (драйверов), полученных из коллекционного центра в Блюмингтоне (США), в которых встроенные векторные конструкции содержат последовательности, кодирующие дрожжевой транскрипционный фактор GAL4 под управлением специфических промоторов, обуславливающих экспрессию в нервной системе. Линии w*; P{GAL4-elav.L}3 и P{w(+mW.hs)=GawB}elav(C155) w(1118); P{w(+mC)=UAS-Dcr- 2.D}2 обуславливают экспрессию в нервной системе в течение всей жизни. Линия w*; P{GawB}D42 обуславливает экспрессию в мотонейронах. Линия w1118; P{Ddc- GAL4.L}4.36 обуславливает экспрессию в дофаминэргических и серотонинэргичес- ких нейронах. Линия w(1118); P{GawB}DJ847 обуславливает уменьшающуюся с возрастом экспрессию в клетках нервной системы. Линия w(1118); P{GawB}DJ651 обуславливает увеличивающуюся с возрастом экспрессию в клетках нервной системы.
Таким образом, в результате проведенной работы появилась возможность увеличивать уровень экспрессии четырех различных форм GSK-3 P (PA, PB, PG и PO) в нервной системе модельных особей, совмещающих в своем геноме дополнительную копию определенного варианта гена shaggy (трансген) и специфический для нервной системы драйвер. Следующий этап работы заключался в характеристике модельных особей. Такая характеристика включала формальный этап - подтверждение того, что у модельных особей действительно увеличено количество транскрипта, соответствующего определенной форме GSK-3 в, и содержательный этап - подтверждение того, что модельные особи имеют фенотипические изменения, позволяющие судить о наличии у них признаков таупатии: а) появление агрегированного (гиперфосфорилированного) тау; б) нарушение функции синапсов; в) изменение поведения и памяти; г) изменение подвижности; д) изменение продолжительности жизни. Далее будут представлены предварительные характеристики модельных особей с генотипами w(1118)/P{w(+mW.hs)=GawB}elav(C155) w(1118); +/P{w(+mC)=UAS-Dcr-2.D}2; P{w(+mC)=UAS-sgg.B}MB5/+, и w(1118)/P{w(+mW hs)=GawB}elav(C155) w(1118); +/P{w(+mC)=UAS-Dcr-2.D}2; P{w(+mC)=UAS-sgg. Y214F}2/+, у которых сильно и умеренно увеличена экспрессия основной формы GSK-3 в, PB, в клетках нервной системы на протяжении всей жизни, по сравнению с контрольными особями с генотипом w(1118)/P{w(+mW.hs)=GawB}elav(C155) w(1118); +/P{w(+mC)=UAS-Dcr-2.D}2; +/+.
Для того чтобы понять, действительно ли индукция трансгена приводит к сверхэкспрессии основной формы GSK-3 в, необходимо было измерить количество транскрипта, соответствующего форме RB, после индукции. Из самцов и самок модельных и контрольного генотипов на стадии личинки третьего возраста и на стадии имаго в возрасте 1-3 дней была выделена РНК, получена кДНК, и количество транскрипта RB было измерено с помощью ПЦР в реальном времени. Для амплификации были использованы праймеры, специфичные для данной формы транскрипта. И у личинок, и у взрослых мух обоего пола количество транскрипта RB у особей двух модельных генотипов превышало его количество у контрольных особей (рис. 3 иллюстрирует этот факт на примере личинок самцов и самок модельного генотипа со сверхэкспрессией нормальной основной формы GSK-3 в). Таким образом, на уровне транскрипта у модельных особей действительно наблюдается увеличенная экспрессия гена shaggy, кодирующего GSK-3 в. Следует отметить, что увеличение количества транскрипта сходно у особей двух модельных генотипов; этот результат ожидаем, поскольку экспрессия трансгена в обоих случаях находится под регуляцией одной и той же промоторной последовательности UAS. Мы предполагаем, что количество и суммарная активность белка GSK-3 в PB у особей двух модельных генотипов также увеличены, хотя и в разной степени, что обусловлено свойствами трансгенов, несущих нормальную и мутантную последовательности гена shaggy. Это предположение нуждается в дальнейшей экспериментальной проверке.
Для характеристики функционального состояния синапсов было проанализировано распределение и количество флюоресцентно-меченого маркера активных синаптических зон, белка Bruchpoint (Matkovic et al., 2013) в синаптических окончаниях в районе четвертого мышечного сегмента личинок III возраста. Сверхэкспрессия нормальной основной формы GSK-3 в привела к существенному уменьше-

Рис. 3. Количество транскрипта RB у личинок со сверхэкспрессией нормальной основной формы GSK-3P в нервной системе
Диаграмма построена с помощью программного обеспечения, поставляемого с детектирующим амплификатором MiniOpticon (Bio-Rad)
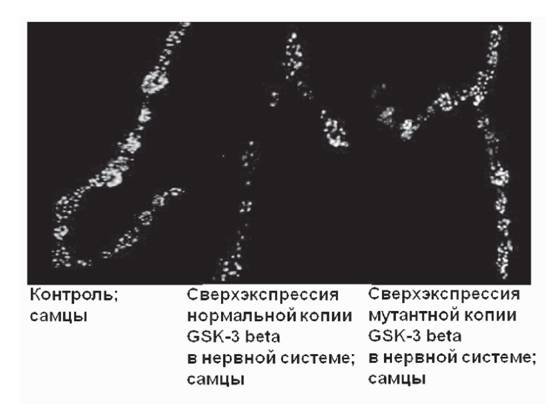
Рис. 4. Активность синапсов у личинок со сверхэкспрессией нормальной и мутантной основной формы GSK-3 в в нервной системе
Иммуногистохимическое окрашивание на белок Bruchpoint (красный)
нию количества активных зон как у самок, так и у самцов (рис. 4), что указывает на нарушение функции синапсов. Сверхэкспрессия мутантной основной формы GSK- 3 P привела к менее выраженным эффектам (см. рис. 4). Исследование влияния сверхэкспресии различных вариантов GSK-3 P на функции нервной системы будет продолжено. Мы планируем охарактеризовать функциональное состояние синапсов в мозге (атенальных долях и грибовидных телах, где присутствуют скопления синапсов) самцов и самок разного возраста; наличие агрегированного (гиперфос- форилированного) тау; уровень нейродегенерации; изменение поведения и памяти у мух модельных генотипов.
У самцов и самок, экспрессирующих основную форму GSK-3 P в нервной системе на протяжении всей жизни, была охарактеризована продолжительность жизни и подвижность. Метод измерения продолжительности жизни описан в (Krementso- va et al., 2012). Для измерения подвижности был использован специальный прибор фирмы TriKinetiks (США). Метод измерения основан на том, что интенсивность передвижения мух в тестерной пробирке оценивается по числу пересечений ими инфракрасных лучей, которое фиксируется компьютером с помощью специальной программы.
Сверхэкспрессия нормальной основной формы GSK-3 P драматически снизила продолжительность жизни и подвижность как самцов, так и самок. Самцы оказались практически неспособными вылезать из куколок и гибли. Если их удавалось вытащить, они оказывались жизнеспособными, однако медианная продолжительность их жизни составила 2 дня, а максимальная не превышала 4 дней, при медианной продолжительности жизни контрольных самцов 74 дня и максимальной - 92 дня. Медианная продолжительность жизни модельных самок составила 4 дня, а максимальная - 22 дня (77 и 96 дней у контрольных самок). Подвижность таких самцов и самок была нарушена настолько, что плохо детектировалась прибором. Сверхэкспрессия мутантной основной формы GSK-3 P снизила продолжительность жизни самцов (медианная продолжительность жизни - 42 дня, максимальная - 74 дня) и практически не изменила продолжительность жизни самок (медианная продолжительность жизни - 71 день, максимальная - 84 дня). Подвижность самцов и самок не отличалась от нормальной. Таким образом, степень изменения продолжительности жизни определяется как трансгеном, так и полом особей. Предполагаемое сильное увеличение активности GSK-3 P существенно уменьшает продолжительность жизни, в то время как предполагаемое умеренное увеличение активности GSK-3 P уменьшает продолжительность жизни в меньшей степени. И в том, и в другом случае более сильная реакция наблюдается у самцов.
Таким образом, первые данные о свойствах модельных особей позволяют предположить, что на их основе может быть успешно создана модель таупатии. Различные варианты модели позволяют исследовать различные характеристики таупатии. Так, у модельных особей со сверхэкспрессий мутантной основной формы GSK-3 Р, характеризующихся нормальной жизнеспособностью и продолжительностью жизни, можно исследовать зависимые от возраста поведение и память мух. Модель, основанная на сверхэкспрессии нормальной основной формы GSK-3 Р, характеризующаяся более выраженными эффектами в отношении синаптической функции, может быть использована для дальнейшего изучения патологических изменений нервной системы. Мы также предполагаем провести исследование Drosophila me- lanogaster со сверхэкспрессией других форм GSK-3 р. Такой подход позволит всесторонне оценить роль GSK-3 P в контроле продолжительности жизни и функции нервной системы, включая поведение и память, выявить молекулярные основы взаимосвязи между этими признаками в норме и патологии.
Литература
Agrawal N., Pallos J., Slepko N. et al. Identification of combinatorial drug regiments for treatment of Huntington’s disease using Drosophila // Proc. Natl. Acad. Sci. USA. 2005. Vol. 102. P. 3777-3781.
Ataman B., Ashley J., Gorczyca M. et al. Rapid activity-dependent modifications in synaptic structure and function require bidirectional Wnt signaling // Neuron. 2008. Vol. 57. P. 705-718.
BourouisM. Targeted increase in shaggy activity levels blocks Wingless signaling // Genesis. 2002. Vol. 34. P. 99-102.
Buerger K., Ewers M., Pirttila T., et al.CSF phosphorylated tau protein correlates with neocortical neurofibrillary pathology in Alzheimer’s disease // Brain. 2006. Vol. 129. Pt. 11. P. 3035-3041.
Cauchi R.J., van den Heuvel M. The fly as a model of neurodegenerative diseases: is it worth the jump? // Neurodegenerative Dis. 2006. Vol. 3. P. 338-356.
Chen X., Li Y., Huang J. et al. Study of tauopathies by comparing Drosophila and human tau in Drosophila // Cell. Tissue Res. 2007. Vol. 329. P. 169-178.
Chiang A., Priya R., Ramaswami M., et al. Neuronal activity and Wnt signaling act through Gsk3-P to regulate axonal integrity in mature Drosophila olfactory sensory neurons // Development. 2009. V 136. P. 1273-1282.
Compta Y., Parkkinen L., Kempster P. et al. The significance of a-synuclein, amyloid-P and tau pathologies in Parkinson's disease progression and related dementia // Neurodegener. Dis. 2013.doi: 10.1159/000354670.
Dionne VS., Schneider D.S. Models of infectious diseases in the fruit fly Drosophila melanogaster // Disease Models Mech. 2008. Vol. 1. P. 43-49.
Froldi F., Ziosi M., Tomba G. et al. Drosophila Lethal Giant Larvae neoplastic mutant as a genetic tool for cancer modeling // Current Genomics. 2008. Vol. 9. P. 147-154.
Jackson G.R., Wiedau-Pazos M., Sang T. K. et al. Human wild-type tau interacts with wingless pathway components and produces neurofibrillary pathology in Drosophila // Neuron. 2002. Vol. 34. P. 509-519.
Jeibmann A., Paulus W. Drosophila melanogaster as a model organism of brain diseases // Int. J. Mol. Sci. 2009. Vol. 10. P. 407-440.
Kapahi P., Chen D., Rogers A.N. et al. With TOR, less is more: a key role for the conserved nutrient-sensing TOR pathway in aging // Cell Metab. 2010. Vol. 11. P. 453-465.
Kawakami F., Shimada N., Ohta E. et al. LRRK2 regulates tau phosphorylation through direct activation of GSK-3P // FEBS J. 2013. doi: 10.1111/febs.12579.
Konsolaki M. Fruitful research: drug target discovery for neurodegenerative diseases in Drosophila // Expert. Opin.Drug Discov. 2013. Vol. 8. P. 1503-1513.
Krementsova A.V., Roshina N. V., Tsybul’ko E.A. et al. Reproducible effects of the mitochondria-targeted plastoquinone derivative SkQ1 on Drosophila melanogaster lifespan under different experimental scenarios // Biogerontology. 2012. Vol. 13. P. 595-607.
Lai T. S., Liu Y., Tucker T. et al. Identification of chemical inhibitors to human tissue transglutaminase by screening existing drug libraries // Chem. Biol. 2008. Vol. 15. P. 969-978.
Lu B., Vogel H. Drosophila models of neurodegenerative diseases // Annu. Rev. Pathol. Mech. Dis. 2009. Vol. 4. P. 315-342.
Mal ’tsev A.V.,Davidchenko N.V., Uteshev V.K. et al. Intensive protein synthesis in neurons and phosphorylation of beta-amyloid precursor protein and tau-protein are triggering factors of neuronal amyloidosis and
Alzheimer’s disease // Biomed. Khim. 2013. Vol. 59. P. 144-170.
Matkovic T., SiebertM., Knoche E. et al. The Bruchpilot cytomatrix determines the size of the readily releasable pool of synaptic vesicles // J. Cell Biol. 2013. Vol. 202.P. 667-683.
Medina M., Avila J. New insights into the role of glycogen synthase kinase-3 in Alzheimer’s disease // Expert Opin. Ther. Targets. 2014. Vol. 18. P. 69-77.
Mershin A.,Pavlopoulos E., Fitch O. et al. Learning and memory deficits upon TAU accumulation in Drosophila mushroom body neurons // Learn. Mem. 2004. Vol. 11. P. 277-287.
Mudher A., Shepherd D., Newman T.A. et al. GSK-3beta inhibition reverses axonal transport defects and behavioural phenotypes in Drosophila // Mol. Psychiatry. 2004. Vol. 9. P. 522-530.
Nehme N.T.,Lie'geois S., Kele B. et al. A model of bacterial intestinal infections in Drosophila melanogaster // PLoS Pathol. 2007. Vol. 3. P. e173.
Noble W., Hanger D. P., Miller C. C., Lovestone S. The importance of tau phosphorylation for neurodegenerative diseases // Front. Neurol. 2013. Vol. 4. P. 83.
Palomero T., DominguezM., Ferrando A.A. The role of the PTEN/AKT pathway in NOTCH1-induced leukemia // Cell Cycle. 2008. Vol. 7. P. 965-970.
Pan X.D., Chen X.C. Clinic, neuropathology and molecular genetics of frontotemporal dementia: a minireview // Transl. Neurodegener. 2013. Vol. 2. P. 8-14.
Pandey U.B., Nichols C.D. Human disease models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery // Pharmacol. Rev. 2011. Vol. 63. P. 411-436.
Pooler A.M.,Polydoro M., Wegmann S. et al. Propagation of tau pathology in Alzheimer’s disease: identification of novel therapeutic targets // Alzheimers Res. Ther. 2013. Vol. 5. P. 49-61.
Ramachandran G., Udgaonkar J.B. Mechanistic studies unravel the complexity inherent in tau aggregation leading to Alzheimer’s disease and the tauopathies // Biochemistry. 2013. V.52. P. 4107-4126.
ReadR.D., Cavenee W.K., FurnariF.B., Thomas B.A. Drosophila model for EGFR-Ras and PI3K-dependent human glioma // PLoS Genet. 2009. Vol. 5. P. e1000374.
Reddy P.H. Amyloid beta-induced glycogen synthase kinase 3p phosphorylated VDAC1 in Alzheimer's disease: implications for synaptic dysfunction and neuronal damage // Biochim. Biophys. Acta. 2013. Vol. 1832. P. 1913-1921.
Rockenstein E., Crews L., Masliah E. Transgenic animal models of neurodegenerative diseases and their application to treatment development // Adv. Drug Deliv. Rev. 2007. Vol. 59. P. 1093-1102.
Roeder T.,Isermann K., KabeschM. Drosophila in Asthma Research // Am. J. Respir. Crit. Care. Med. 2009. Vol. 179. P. 979-983.
Rowe M.K.,Wiest C., Chuang D.M. GSK-3 is a viable potential target for therapeutic intervention in bipolar disorder // Neurosci. Biobehav. Rev. 2007. Vol. 31. P 920-931.
Shcherbata H.R.,Yatsenko A.S., Patterson L. et al. Dissecting muscle and neuronal disorders in a Drosophila model of muscular dystrophy // EMBO J. 2007. V 26. P. 481-493.
Sibley C.D., Duan K., Fischer C. et al. Discerning the complexity of communityinteractions using a Drosophila model of polymicrobial infections // PLoS Pathol. 2008. Vol. 4. P. e1000184.
Sofola O., Kerr F., Rogers I. et al. Inhibition of GSK-3 ameliorates Abeta pathology in an adult-onset Drosophila model of Alzheimer‘s disease // PLoS Genet. 2010. Vol. 6. P. e1001087.
Taghli-Lamallem O., Akasaka T., Hogg G. et al. Dystrophin deficiency in Drosophila reduces lifespan and causes a dilated cardiomyopathy phenotype // Aging Cell. 2008. Vol. 7. P. 237-249.
WilliamsD.W.,TyrerM., ShepherdD. Tau and tau reporters disrupt central projections of sensory neurons in Drosophila // J. Comp. Neurol. 2000. V. 428. P. 630-640.
Wittmann C.W., Wszolek M.F., Shulman J.M. et al. Tauopathy in Drosophila: neurodegeneration without neurofibrillary tangles // Science. 2001. Vol. 293. P. 711-714.
Wolf M.J., Amrein H., Izatt J.A. et al. Drosophila as a model for the identification of genes causing adult human heart disease // Proc. Natl. Acad. Sci. USA. 2006. Vol. 103. P. 1394-1399.
Xu Y., Condell M., Plesken H. et al. A Drosophila model of Barth syndrome // Proc. Natl. Acad. Sci. USA. 2006. Vol. 103. P. 11584-11588.
Yeh PA., Chien J.Y., Chou C.C. et al. Drosophila notal bristle as a novel assessment tool for pathogenic study of Tau toxicity and screening of therapeutic compounds // Biochem. Biophys. Res. Commun. 2010. Vol. 391. P. 510-516.
Еще по теме Молекулярно-генетическая модель таупатии на основе сверхэкспрессии протеинкиназы GSK3p в нервной системе Drosophila melanogaster:
- Drosophila melanogaster - модельная система для изучения потенциальных терапевтических соединений
- Моделирование патогенеза нейродегенеративных заболеваний на Drosophila melanogaster
- Преимущества Drosophila melanogaster для моделирования нейродегенеративных заболеваний человека
- Выбор терапии у больных раком желудка на основе молекулярно-генетических маркеров
- Перспективы разработки превентивного лечения болезни Паркинсона на основе мобилизации системы молекулярных шаперонов
- Молекулярно-генетическое исследование
- Молекулярно-генетическое исследование
- Молекулярно-генетические маркеры.
- Молекулярно генетические аспекты канцерогенеза при раке желудка
- Молекулярно-генетические методы исследования
- Молекулярно-генетические болезни.
- Молекулярно-генетические маркеры, изучаемые в работе
- Глава 5. Молекулярный скрининг генетического риска.
- Экспериментальная проверка математических моделей прогнозирования и ранней диагностики заболеваний нервной и иммунной системы в условиях действия электромагнитных полей.
- Генетические модели