9.0.4. Простые модели ферментов
Все известные ферменты — это белки. Однако для изучения механизмов их действия и установления природы их высокой специфичности в отношении и продуктов и субстратов были созданы модельные соединения небелковой природы.
Основой для них послужил гистидин, встречающийся в активных центрах многих ферментов. Наиболее важной функциональной группой гистидина является имидазольное кольцо (9.1), величина рКа которого близка 7. Это позволяет ему выступать в роли как донора протона, так и его акцептора. Имидазол и особенно 4'-(имидазол-4-ил) масляная кислота (9.2) значительно ускоряют гидролиз сложных эфиров по механизму обычного кислотно-основного катализа [Bruice, Benkovic, 1966].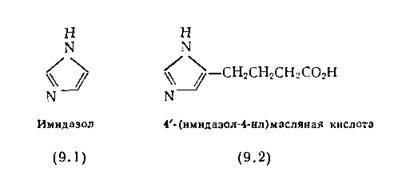
По свойствам наиболее близки ферментам циклодекстрины (циклоамилазы), получаемые при расщеплении крахмала [Cramer, 1956]. Циклодекстрины действуют как эффективные эстеразы, субстратами которых служат фенилацетат и его производные. По данным рентгеноструктурного анализа молекулы циклодекстринов представляют собой тор из шести остатков a-D-глюкозы в обычной конформации. Гидроксильные группы образуют короны вокруг верхней и нижней части тора, внутренний диаметр которого составляет около 0,5 нм. При гидролизе карбонильная группа ацетата («гость») приближается к вторичным гидроксильным группам циклодекстрина («хозяин»): чем теснее этот контакт, тем быстрее проходит гидролиз. Сложный эфир («гость») ацилирует одну из гидроксильных групп «хозяина», высвобождая при этом фенол. Затем в результате реакции внутримолекулярного катализа происходит регенерация циклодекстрина в неацилированную форму. Процесс при этом может повторяться неограниченно. Этот цикл реакций напоминает ферментативные превращения еще и тем, что он подчиняется кинетике Михаэлиса — Ментен (разд. 9.1) и легко ингибируется нормальными субстратами [Bender, Komiyama, 1978].
Ковалентным присоединением двух имидазольных групп к циклодекстрину можно получить искусственную рибонуклеазу, имеющую достаточно высокую специфичность в отношении как субстрата, так и продукта [Breslow, 1983]. Присоединяя ковалентно к циклодекстрину пиридоксамин, можно получить синтетическую трансаминазу. Эти модельные соединения вызывают ускорение соответствующих реакций в 200 раз, однако в ближайшем будущем можно надеяться на значительно большее увеличение скорости [Breslow, 1983].
Исследования этих модельных соединений и данные рентгеноструктурного анализа природных ферментов показали, что для гидролиза необходимы сближение нуклеофильной группы (аниона глутаминовой кислоты, аспарагиновой кислоты или имидазола) с электрофильным центром — атомом углерода карбонила амидной группы и одновременный подход протона к атому азота этой же группы. На циклодекстриновых моделях было установлено, что ферментативноподобная активность может быть свойственна веществам с ОММ несколько ниже 1000, имеющим простую структуру и симметрию. Более подробные сведения о моделях ферментов можно получить из работ Jencks (1969) и Bender, Komiyama (1978). В литературе есть данные о синтезе микроцикла, ускоряющего ацетилирование аминокислот в 1011 раз [Cram, 1983].
Действие ферментов в значительной степени зависит от мицелл амфифильных молекул, катализирующих многие химические реакции и нередко рассматриваемых как очень простые модели ферментов (определение понятий «амфифильный» и «мицелла» приведено в разд. 14.0). Расположение гидрофобных (внутри) и гидрофильных (снаружи) групп в мицелле напоминает расположение этих групп в молекуле фермента. Подобно ферментам мицеллы денатурируют нагреванием или действием мочевины; как и ферменты, они обладают избирательностью по отношению к субстратам [Jencks, 1969]. Реакции, сопровождающиеся высвобождением анионов, такие как гидролиз сложных эфиров, катализируются катионными мицеллами, например мицеллами цетилтриметиламмоний бромида (14, 16) [Menger, Portnoy, 1967]. Субстрат концентрируется в мицелле. При этом образуются ван-дер-ваальсовы связи, о чем свидетельствует повышение эффективности катализа при увеличении размеров молекул субстрата. Заряженные группы мицеллы создают заряд в среде, окружающей субстрат (это окружение может стабилизировать переходное состояние субстрата); предполагается, что фактически они и являются катализаторами. Низкая диэлектрическая постоянная остальной части мицеллы может приводить к дестабилизации любого заряда на субстрате, его переходном состоянии и продуктах.
Эффективность мицеллярного катализа иллюстрирует доде- цилсульфат натрия, ускоряющий связывание ионов Си2+ пор- фиринами в 500 000 раз [Lowe, Phillips, 1961]. О катализе мицеллами см. Cordes (1973), Fendler и Fendler (1975).
Еще по теме 9.0.4. Простые модели ферментов:
- Ферменты пищеварительного тракта
- Ферменты пищеварительного тракта
- 9.0.5. Биологическая регуляция ферментов
- 9.0. Ферменты
- 9.0.1. Конформационная адаптация фермента к субстрату
- 9.0.6. Ферменты и лекарственные вещества
- Амилоид-деградирующие ферменты
- Тип Protozoa — простейшие
- Простое окрашивание
- Простой герпес
- Простейшие (общая характеристика)
- Культивирование простейших кишечника
- Необратимые ингибиторы активного центра ферментов (НИАЦФ)
- Ферменты процессинга Ар
- Регуляция свойств амилоид-деградирующих ферментов
- Просто гигиена.