Свободнорадикальное окисление липидов в тканях глаза
B последние годы очень большое внимание уделяется изучению ПОЛ (Владимиров Ю.А. и др., 1972, 1975; Воскресенская Л.К., 1984; Эфендиев H.M., 1972, 1978, 1992; Эфендиев H.M. и др., 1985a,6, 1989, 1991, 1995; Эмануэль H.M.
и др., 1964, 1968).ПОЛ является чрезвычайно сложным процессом и включает в себя несколько элементарных реакций, которые можно разбить на 4 стадии:
1) инициирование цепи:
K1
RH+X' —> R'+XH;
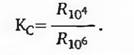
B тканях хорошо сбалансированы реакции образования и расходования продуктов ПОЛ и они протекают на низком стационарном уровне. При развитии различных патологических состояний тканей баланс может нарушиться, образующиеся гидроперекиси будут накапливаться, приводя к серьезным нарушениям в биологических мембранах (Владимиров Ю.А. и др., 1972; Воскресенский O.H., 1975; Джафаров А.И., 1976, 1982; Джафаров А.И. и др., 1985; Кулиева Э.М., 1982; Коган B.E. и др., 1973, 1979, 1981, 1985). При этом низкий уровень перекисей липидов, свойственный нормальным тканям, совсем не означает, что в них не происходит ПОЛ.
Изучение ПОЛ в организме выявило участие продуктов ПОЛ в развитии многих патологических состояний и заболеваний.
Протекторную роль антиоксидантов наблюдали при консервировании и длительном хранении, катарактогенезе, диабетической ретинопатии, витреохориоретинальной дистрофии СДжафаров А.И., 1980; Ates N.A. et al., 2004; Babizhayeva M. et al., 2004; Di Leo M.A. et al., 2003; Ertekin M.V. et al., 2004; Galkina O.V. et al., 2003; Ka- razhaeva МЛ. et al., 2004; Kozlov S.A. et al., 2003; Obrosova G. et al., 2001; Tanun B.B. et al., 2004; Wachtlin J. et al., 2000).
O возможности коррекции интенсификации ПОЛ в тканях глаза при ожогах разными природными и синтетическими антиоксидантами указывали А.И.Еременко и др. (1994), И.П.Метелицина и др.
(2002), J.L.Alio et al. (1995), E.Banin et al (2003), S.J.Till et al. (2003), A.G.Travkin et al. (2004), H.P.Yuan et aL (1994).Установлена интенсификация ПОЛ в тканях глаза при экспериментальном офтальмогерпесе. По мнению Б.С.Касавиной и соавт. (1982), ПОЛ является пусковым механизмом для развития данного патологического процесса. B результате ПОЛ содержание МДА в острый период заболевания в ткани роговицы возрастает в 7 раз, во влаге передней камеры — в 3.5 раза, а в радужной оболочке — в 2.8 раза.
Интенсификации ПОЛ в ткани роговицы при офтальмогерпесе может способствовать снижение в ней активности антиоксидантов. Так, активность глутатионпероксидазы (ГП) в динамике инфицирования роговицы и других тканей глаза вирусом простого герпеса в роговице в несколько раз меньше, чем в радужке или в хрусталике (Алиева З.А. и др., 1958).
Данные экспериментально-клинических исследований (Kaca- вина Б.С. и др., 1982; Herr R.D. et al., 1991) свидетельствуют о том, что инстилляция 10% а-токоферола (витамин E) снижает содержание малонового диальдегида (МДА), повышает активность антиоксидантов, что способствует уменьшению количества осложнений и неприятных последствий герпетического поражения глаз, сокращает длительность стационарного лечения больных офтальмогерпесом.
Изменение интенсивности хемилюминесценции в ткани роговицы при ее консервации доказывает, что в фосфолипидах клеточных мембран создаются условия для накопления продуктов свободнорадикального окисления (CPO), приводящего к разрушению биологических структур, что может отрицательно влиять на приживление трансплантата (Федоров C.H. и др., 1984).
Возможность активации ПОЛ в водянистой влаге и омывающих ее тканевых структурах глаза обусловлена генерацией и накоплением в камерной влаге H2O2- H2O2, содержащаяся в водянистой влаге, может при определенной концентрации продуктов CPO во влаге передней камеры повреждать не только хрусталик, но и трабекулу и стенки шлеммового канала (Владимиров Ю.А. и др., 1972; Воскресенская Л.К., 1984; Журавлев А.И., 1975, 1988).
Этот факт позволяет предположить участие ПОЛ в развитии деструктивных изменений при открытоугольной глаукоме и объясняет гипотензивное действие инстилляций и субконъюнктивальных введений 4% раствора тауфона, являющегося естественным антиоксидантом, при повышении ВГД в результате резкого увеличения секреции водянистой влаги и его нормализующее действие на нарушенную проницаемость гематоофтальмического барьера. Имеются сведения об интенсификации ПОЛ и возможном участии его продуктов в патогенезе катаракты (Лазаренко Л.Ф. и др., 1986).Мы наблюдали повышение уровня продуктов ПОЛ у больных катарактой, а также при одной из разновидностей наследственной катаракты у экспериментальных животных. Инициация ПОЛ, по нашему мнению, происходит за счет активных форм кислорода и продуктов фотоокисления триптофана — производных кинуренина.
Установлено, что при катарактах разной этиологии снижается активность супероксиддисмутазы (СОД), ГП, глутатионтрансфе- разы, глутатиона и аскорбиновой кислоты (Воскресенская Л.К., 1984; Воскресенский O.H., 1975).
Еще в 1985 г. З.А.Алиева и соавт. в сравнительном аспекте изучили содержание компонента антиоксидантной системы — восстановленного глутатиона и активность ГП в тканях глаза быка и человека и выявили, что наивысшим уровень указанных антиоксидантов был в роговице и хрусталике.
Н.М.Эфендиев с соавт. (1972, 1978, 1985a,6, 1987a,6, 1988, 1989, 1991, 1995) выделили 3 универсальных механизма повреждения мембранных структур, которые могут играть важную роль в старческом катарактогенезе:
• активация ПОЛ;
• накопление липидов и увеличение активности фосфолипаз;
• накопление продуктов деградации фосфолипидов, разрушающих мембраны.
При изучении влияния света на водорастворимые белки хрусталика В.Е.Охрименко (1962) установлено, что в реакциях фото- сенсибилизированного окисления белков хрусталика участвует синглетный кислород, присутствие которого может быть одной из причин катаракты.
Имеются сведения об интенсификации ПОЛ и об участии его продуктов при увеитах, осложненной миопии, анафилактическом шоке, эндофтальмитах, аутолитическом увеите экспериментальных животных (Каримова M.X.
и др., 2002; Катаргина Л.А. и др., 2003). Введение антиоксидантов (каталазы, бензоата натрия) обеспечивало сохранность структур глаза, в частности структур сетчатки, уменьшало явления воспалительной экссудации и воспаления в стекловидном теле. Обнаруженный защитный эффект антиоксиданта свидетельствует об участии H2O2, свободнокислородных и гидроксильных радикалов в развитии увеита.Важную роль в возникновении дистрофических заболеваний сетчатки, в частности центральных инволюционных дистрофий сетчатки, играет интенсификация ПОЛ и увеличение его продуктов (Эфендиев H.M. и др., 1972, 1978, 1985a,6, 1987a,6, 1988, 1989, 1991, 1995).
Положительный результат отмечен при лечении пигментной дистрофии сетчатки антиоксидантом тауфоном.
Интенсификация ПОЛ имеет место при интравитриальных кровоизлияниях различной этиологии (Нейман-Заде H.K. и др., 1987a,6). Авторами установлено, что между уровнем ПОЛ, в частности, содержанием МДА и интенсивностью хемилюминесценции, существует определенный параллелизм. Наибольший уровень CPO наблюдался у больных гемофтальмом с диабетической этиологией. При этом уровень спонтанной и индуцированной хемилюминесценции стекловидного тела превышал контрольный в 9.8 раза (Нейман-Заде H.K. и др., 1987a). При экспериментальном гемофтальме уровень интенсивности хемилюминесценции сетчатки увеличивался, начиная с 6-х суток. B течение 2 мес уровень CPO непрерывно возрастал, к концу 2-го месяца он увеличился в 4-6 раз, а к исходу 3-5-го месяца уровень метаболического свечения сохранялся прежним.
По мнению Н.К.Нейман-Заде и соавт. (1987a), интенсификация ПОЛ в сетчатке в результате интравитриальных кровоизлияний усугубляет перекисное повреждение в стекловидном теле, что может иметь важное значение в швартообразовании.
Таким образом, возникает своего рода замкнутый “порочный круг”, превращающий стекловидное тело в “депо” токсических веществ.
Интравитриальные кровоизлияния как у людей, так и у экспериментальных животных сопровождаются заметной активацией ПОЛ, скорость и характер развития которого во многом зависят от количества излившейся крови, локализации кровоизлияния и состояния стекловидного тела.
По данным Н.М.Эфендиева с соавт. (19876, 1991, 1988a,6, 1989), при интравитриальных кровоизлияниях в стекловидном теле создаются условия, которые могут способствовать усилению ПОЛ, в частности, усиление генерации радикальных интермедиатов кислорода (*O2-, OH*), изменение липидного состава стекловидного тела, увеличение концентрации металлов переменной валентности (Fe, Cu), наличие свободного гемоглобина. Особое значение при этом имеют усиленные образования радикальных интермедиатов кислорода и вносимые в стекловидное тело в составе аутокрови ионы металлов переменной валентности и гемоглобин (внутри- и внеэритроцитарный).
Железо регулирует скорость ПОЛ в тканях. При этом каталитическом действии происходит распад H2O2 с образованием свободных радикалов HO* и HO-, обладающих наиболее повреждающим действием (Владимиров Ю.А. и др., 1972, 1975; Воскресенская Л.К., 1984). B митохондриях или суспензиях фосфолипидов без добавления ионов железа извне H2O2 не накапливается. B то же время процесс инициирования окисления жирных кислот протекал в присутствии Fe3+ или при добавлении дериватов гемоглобина, в частности геминсодержащих соединений (Владимиров Ю.А. и др., 1972).
Измерение интенсивности хемилюминесценции при определении относительной эффективности радикалообразования из H2O2 при разных концентрациях гемоглобина, гемина и ионов железа позволило Н.К.Нейман-Заде (1988) установить, что внеэритроцитар- ный гемоглобин и продукты его деструкции, вплоть до гемина, являются наиболее эффективными прооксидантами.
Изучая роль каждой из систем, участвующих в процессе интенсификации ПОЛ при экспериментальных кровоизлияниях в стекловидное тело, Н.М.Эфендиев с соавт. (1976, 1985a,6, 1987a,6, 1991, 1988, 1989) выявили, что почти идентичная кинетика в накоплении продуктов ПОЛ и изменение интенсивности хемилюминесценции, наблюдаемые при введении аутокрови, воспроизводятся при комбинированном введении в стекловидное тело железа и меди с гемоглобином и эмульсией фосфолипидов. При введении этих ингредиентов по отдельности величины показателей ПОЛ были иными.
Наиболее быстрое действие оказывали гемоглобин и железо.Интенсификация ПОЛ с участием ионов металлов переменной валентности при интравитриальных кровоизлияниях может способствовать развитию структурно-функциональных нарушений в стекловидном теле и сетчатке. Так, при экспериментальных кровоизлияниях в стекловидное тело радикалы *OH, генерированные H2O2, вызывают неферментативную деполимеризацию витриального геля при каталическом действии ионов железа из разрушенного гемоглобина или от металлических осколков, приводя к разжижению стекловидного тела.
По данным Н.М.Эфендиева, железо также может катализировать образование *OH и HO2 и без участия супероксиданион-ради- калов.
Данные Н.М.Эфендиева свидетельствуют о способности кислородных радикалов, порожденных разными системами генерации, деполимеризовать гиалуроновую кислоту. Так, поток кислородных радикалов приводит к изменению размеров (появление полидисперсности) и конформационной характеристики (понижение вязкости) полисахарида.
Н.К.Нейман-Заде с соавт. (1987a,6) обнаружили, что энзима- тически образованный супероксиданион-радикал (*O2) вступает в реакцию с H2O2, образуя гидроксилрадикал (OH*) и тем самым вызывая деполимеризацию гиалуроновой кислоты. По их мнению, для деполимеризации гиалуроновой кислоты присутствие H2O2 и O2 является необходимым. Косвенным подтверждением этому служат экспериментальные данные Н.К.Нейман-Заде с соавт., свидетельствующие об участии H2O2 в ингибировании синтеза гиалуроновой кислоты в результате лечения гипоксантином.
К.Т.Керимов с соавт. (2000) в экспериментах показал, что синглетный кислород ('O2) вызывает изменения в структуре гиалуроновой кислоты, что выражается в понижении относительной вязкости полисахарида. Предположительно, понижение относительной вязкости связано как с уменьшением молекулярного веса, так и с изменением структуры макромолекулы гиалуроновой кислоты, и возникает в результате воздействия синглетного кислорода, причем относительная вязкость постоянно уменьшается при увеличении продолжительности действия 'O2.
Накопление продуктов ПОЛ наряду с изменением вязкости стекловидного тела может вызывать полимеризацию его белков, в частности, поперечную сшивку одного из структурных компонентов-------------------------------------------------------
ацетилглюкозамина, вследствие чего могут образовываться шварты (Эфендиев H.M. и др., 1976, 1985a,6, 1987a,6, 1991, 1988, 1989).
Н.М.Эфендиев (1992) с помощью морфологических и биофизических исследований тканей в разные сроки экспериментального гемофтальма показал, что в далеко зашедших случаях швартооб- разования резко усиливалась интенсивность фотохемилюминесценции, указывающей на накопление свободнорадикальных продуктов, которая приводила к резкому нарушению метаболизма ткани. Накопление свободнорадикальных продуктов, по мнению автора, играет большую роль в формировании белковых образований, из которых образуется соединительнотканный рубец.
Электронномикроскопическими и гистохимическими исследованиями тканей при экспериментальном гемофтальме установлено, что токсическое действие на сетчатку ионы железа начинают оказывать на ранних этапах кровоизлияния, о чем свидетельствует присутствие на 7-е сутки после введения аутокрови железосодержащих комплексов в межклеточных пространствах фоторецепторных клеток сетчатки (Багров C.H., 1977; Багров C.H. и др., 1989).
Наряду с кровью сильное токсическое действие на сетчатку оказывают продукты ПОЛ, образованные в стекловидном теле при интравитриальных кровоизлияниях (Нейман-Заде H.K., 1988). Между тем сетчатка в норме довольно хорошо защищена от токсического действия продуктов CPO. Показан, например, необычно высокий уровень в ней а-токоферола, антирадикальная активность которого хорошо известна. Его концентрация в наружных сегментах достигает 0.1 моль на моль радоксина. Важную протекторную роль в фоторецепторах сетчатки выполняет также система ферментативной защиты от активных форм O2 и его продуктов, которая включает ферменты СОД, глутатионредуктазу и ГП (Бурлакова E.B., 1968, 1978; Коган B.E. и др., 1973, 1979, 1981, 1985).
Таким образом, исходя из данных литературы, следует, что при многих заболеваниях глаз интенсифицируется ПОЛ и нарушается структурно-функциональное состояние тканей глаза. Важную роль при этом играют радикальные интермедиаторы O2: суперок- сиданион-радикалы, гидроксильные радикалы и ионы металлов переменной валентности, генерация и концентрация которых резко увеличивается при указанной патологии.
Роль свободнорадикального окисления в патогенезе химических ожогов глаз
Ожоги каждого вида вызывают характерные морфологические и биохимические изменения в глазу. Щелочи, омыляя жиры и жироподобные вещества клеточных мембран, проникают в клетку, изменяют pH, вызывают растворение белков (колликвационный некроз), образуя при этом щелочные альбуминаты и ряд соединений белковой молекулы, а также фенолы, скатолы, индолы, пептоны и другие соединения, обладающие высокой токсичностью).
Ha способность обжигающих веществ быстро внедряться в ткани, вступать в соединение с ними, вызывать некроз, нарушать обменные процессы, глубоко инфильтрировать роговицу указывали А.А.Ахмедов (1994, 1997, 1998a,6), П.А.Бездетко с соавт. (1992), Д.И.Березинская (1950), В.В.Волков с соавт. (19896).
Гистоморфологические исследования показали, что при химических ожогах деструктивные изменения развиваются во всех оболочках глаза. Некроз тканей сопровождается реактивными воспалительными явлениями, при этом наблюдается паралич стенок сосудов, их дилатация, повреждение эпителия, стромы, эндотелия сосудов, краевой петлистой сети, увеального тракта (Войно-Ясенецкий
B.B., 1970; Григорян В.Г., 1956; Груша O.B. и др., 1989, 1993; Гун- дорова P.A., 1995; Дамбите Г.Р. и др., 1977; Давыдов Д.В., 2000; Дьяченко В.П., 1960).
Вследствие повреждения эпителиальных и эндотелиальных мембран при ожогах роговицы повышается способность мукоидов роговицы поглощать воду. Отек и оводнение мукоидов являются причиной деполимеризации роговицы СДугельный P.A., 1961). Так, потеря коллагеном МПС при ожогах роговицы приводит к глубоким нарушениям структуры и функции стромы.
При ожогах глаз щелочью соотношение между МПС и липидами в роговице резко нарушается. Чем больше липидов накапливается в тканях роговицы, тем тяжелее протекает ожоговый процесс (Дугельный P.A., 1961).
Ранее показано, что в базальной мембране эпителия роговицы содержатся липиды, фосфолипиды, холестерин. Строма роговицы богата кислыми МПС и коллагеном (Багров C.H., 1977). B настоящее время установлено, что фосфолипиды, ненасыщенные и жирные кислоты являются одним из основных источников ПОЛ (Владимиров Ю.А. и др., 1972, 1975; Воскресенская Л.К., 1984).
Доказано, что ПОЛ непрерывно протекает в норме во всех тканях живых организмов, т.е. определенный уровень ПОЛ является одним из показателей гомеостаза (Тарусов Б.Н., 1964; Теселкин О.Б. и др., 1996). Длительное и значительное изменение интенсивности ПОЛ как в сторону повышения, так и в сторону уменьшения характерно для патологического состояния живого организма (Тарусов Б.Н., 1970, 1984).
Свободные радикалы постоянно участвуют как в неферментативном, так и в ферментативном CPO. Кроме того, установлено, что в ПОЛ весьма важную роль играют активные короткоживущие радикалы.
B процессе жизнедеятельности клетки непрерывно образуются свободные радикалы. Патологическое нарушение проявляется не как следствие появления свободных радикалов, а как следствие изменения их количественного содержания и активности. Количественные изменения радикалов нарушают равновесие регуляторных систем в организме, что проявляется в качественно новых, несвойственных организму патологических реакциях (Эфендиев H.M. и др., 1985a,6, 1987a,6, 1991, 1988, 1989).
Согласно современным представлениям, если в клетке или мембране образуется свободный радикал из молекулы жира, белка или нуклеиновой кислоты, то первым с ним прореагирует кислород, т.к. скорость взаимодействия со свободными радикалами у кислорода выше, чем у других соединений. При взаимодействии радикала и кислорода образуется соединение, имеющее перекисный радикал, который вступает во взаимодействие с другим радикалом, теряя при этом избыток свободной энергии. Переход из одной фазы в основную сопровождается освобождением избытка энергии в виде излучения кванта света (Владимиров Ю.А. и др., 1972, 1975; Воскресенская Л.К., 1984; Джафаров А.И., 1986; Джафаров А.И. и др., 1971).
Количество рекомбинированных радикалов по отношению к количеству радикалов, вступивших во взаимодействие в нормальных физиологических условиях, постоянно (Воскресенский O.H., 1975). Нарушение этого равновесия приводит к накоплению конечных продуктов CPO, которое оказывает цитотоксическое действие на все системы жизнедеятельности клетки (Елисеев H.T. и др., 1967; Журавлев А.И. и др., 1961).
Реакция окисления, с которой связано ПОЛ, протекает во взаимодействии с другими метаболическими процессами. B нормальных физиологических условиях окисление липидов проходит B строго стационарном режиме.
Универсальной системой, регулирующей ПОЛ, является система тканевых биоантиокислителей — соединений, способных тормозить CPO жиров и жирных кислот. K этой системе относятся адреналин, аскорбиновая кислота, сульфгидрильные соединения, кера- тиноиды, токоферолы (Воскресенский O.H., 1975).
При действии разных повреждающих агентов: высоких и низких температур, радиации, химических веществ, интенсивность CPO возрастает (Елисеев H.T. и др., 1967; Журавлев А.И., 1988).
B результате этого в пораженных тканях глаза нарушаются процессы окисления и восстановления, т.е. нарушается интенсивность ПОЛ.
B доступной литературе нам не удалось найти работ, освещающих особенности изменения уровня ПОЛ и AOA при ожогах глаз в разных стадиях ожоговой болезни. Нет информации и об ингибирующем влиянии на уровень ПОЛ препаратов, стабилизирующих проницаемость клеточных мембран и подавляющих образование продуктов CPO при ожогах глаз.
Мы изучили роль ПОЛ в патогенезе ожоговой болезни глаз, определили его интенсивность в разных стадиях ожоговой болезни и проследили его изменение под влиянием препаратов, обладающих антиоксидантной активностью и способных нормализовать проницаемость клеточных мембран.
Антиокислительный потенциал и ПОЛ тканей глаза кроликов при химических ожогах глаз под влиянием природных антиоксидантов
Многочисленные исследования последних лет свидетельствуют об участии CPO и ПОЛ в патогенезе дистрофических, воспалительных и инфекционных заболеваний. Bo всех тканях и органах CPO и ПОЛ протекают на низком стационарном уровне, что достигается за счёт функционирования согласованной системы ингибирования CPO антиоксидантами. Однако при многих патологических состояниях это равновесие нарушается за счёт активации CPO или ослабления системы антиоксидантной защиты, что приводит в конечном итоге к повреждению тканей (Керимов K.T. и др., 1988, 2000; Марголик E.B., 1958). Исходя из этого разработка методов лечения ожоговой болезни глаз должна быть сосредоточена на углублённом исследовании системы ингибирования свободных радикалов, а также других факторов, вызывающих необратимые дистрофические процессы в тканях глаза.
B последние годы появились данные по исследованию антиокислительной активности (AOA) плазмы и QK при дистрофических заболеваниях сетчатки и открытоугольной глаукоме. Однако работ, посвящённых исследованию антиокислительного потенциала слезы, тканей глаза, плазмы крови, а также определению конечных продуктов CPO при экспериментальной ожоговой болезни глаз и динамики изменения этих показателей под влиянием природных антиоксидантов, нет.
Мы изучили антиокислительный потенциал СЖ, влаги передней камеры, стекловидного тела, сетчатки, плазмы крови и изменение этих показателей под влиянием разных схем антиоксидантной терапии.
B работе использован 41 кролик (82 глаза) породы шиншилла массой 2.5-3.0 кг, в возрасте 5-6 мес. У кроликов на обоих глазах моделировали дозированный по площади и величине щелочной ожог роговицы III степени. Подопытных животных забивали методом воздушной эмболии на 3, 14 и 30-е сутки после ожога, забирали у них СЖ, влагу передней камеры, стекловидное тело, сетчатку.
I серия опытов. B I серии опытов исследовали AOA влаги передней камеры, стекловидного тела, сетчатки, определяли содержание ТБК-активных веществ (МДА) в тканях глаза.
B контрольной группе (6 кроликов, 12 глаз) проводили противовоспалительную традиционную терапию (инстилляции 30%
раствора сульфацил-натрия, 0.25% раствора левомицетина по 2 капли 6 раз в день, аппликации тетрациклиновой мази 2 раза в день в конъюнктивальный свод).
Животным опытной группы (6 кроликов, 12 глаз) на фоне противовоспалительной терапии проводили комплексное лечение:
• 0.02% раствор гистохрома спустя 2 ч после ожога в глаз 6 раз в день;
• 0.02% раствор гистохрома в дозе 0.3 мл под конъюнктиву ежедневно в течение 3 нед;
• 1% раствор гистохрома, растворённый в 10 мл физраствора, в вену уха кролика капельно;
• антиоксидант гинкго билоба в дозе 3.5 мг/кг перорально вместе с кормом;
• диквертин в дозе 2.5 мг/кг перорально в течение 3 нед.
AOA влаги передней камеры, стекловидного тела и сетчатки
в некротической стадии ожоговой болезни у контрольных обожженных глаз кроликов была ниже, чем у интактных (табл. 6.8), а под влиянием антиоксидантной терапии повышалась и была ниже показателей нормы на 5.11±0.78% во влаге передней камеры, на 4.56±0.57% в стекловидном теле и на 1.14±0.47% в сетчатке. Лечение ожогов глаз с помощью природных антиоксидантов приводило к достоверному повышению AOA влаги передней камеры.
B стадии трофических расстройств, когда резко выражены воспалительные и экссудативные процессы, в контрольной группе AOA влаги передней камеры снижалась на 71.84±2.38%, а в опытной — на 29.72i2.53% по сравнению с нормой (табл. 6.9). Лечение химических ожогов глаз III степени с помощью комплексной терапии, в состав которой входили природные антиоксиданты, приводило к достоверному повышению AOA влаги передней камеры. AOA стекловидного тела в контрольной группе уменьшалась на
Таблица 6.8. Суммарная AOA в тканях глаза кролика при химическом ожоге глаз Ill степени в некротической стадии под влиянием природных антиоксидантов (мкМ, М±т)
Примечание. Здесь и в табл. 6.9-6.12 *p
Еще по теме Свободнорадикальное окисление липидов в тканях глаза:
- Перекисное окисление липидов.
- Математические модели прогнозирования повторного инфаркта миокарда по показателям перекисного окисления липидов и антиокислительной активности
- Особенности состояния перекисного окисления липидов и системы антиоксидантной защиты.
- Уровень эндогенной интоксикации и активность процессов свободнорадикального окисления в контрольной группе и группе больных миомой матки
- Результаты исследования перекисного окисления липидов у беременных обследованных групп
- 4.2. Оценка системы перекисного окисления липидов эритроцитов и плазмы крови
- Определение показателей перекисного окисления липидов (ПОЛ)
- Влияние Ка20880-инозина на процессы перекисного окисления липидов у больных раком орофарингеальной области, получавших химиолучевую терапию
- Влияние повреждающих факторов химиолучевой терапии на выраженность процессов перекисного окисления липидов в слизистой оболочке полости рта белых беспородных крыс
- Влияние Na^SSG-инозина на процессы перекисного окисления липидов в слизистой оболочке полости рта белых беспородных крыс с химиолучевым оральным мукозитом
- Свободнорадикальные процессы и злокачественный рост
- Манипуляции на покровных тканях глазного яблока
- 189. Ранения глаза: общее понятие, классификация, причины, прогноз. Непроникающие ранения глаза
- Глава 2. Разработка математической электрофизической модели глаза для исследования гемодинамики переднего отдела глаза методом транспальпебральной реоофтальмографии
- Классификация липидов