Мутационный канцерогенез.
Предположим, что мутагенный фактор вызвал нарушение в гене-репрессоре 1 и активный репрессор 1 не может быть синтезирован. В этом случае растормаживается ген инициатора клеточного деления и начинается репликация ДНК.
Образовавшиеся в результате деления клетки не располагают геном-репрессором 1, вследствие чего клетки продолжают репликацию ДНК и при делении образуется семейство клеток, способных к беспредельному неконтролируемому делению. Очевидно, они и являются опухолевыми клетками.Аналогичным образом к беспредельному клеточному делению могут привести мутации и других генов данной регуляторной системы, например мутация гена, кодирующего инициатор клеточного деления, в результате которой он становится недоступным тормозящему влиянию гена-репрессора 1.
Наряду с мутационным возможен и эпигеномный канцерогенез, который предполагает приобретение здоровой клеткой опухолевых свойств путем воздействия на геном клетки факторов, которые не принадлежат к геному данной клетки и не вызывают мутацию, но создают устойчивое нарушение нормальной регуляции генома, приводящее к беспредельному росту
Передающееся из поколения в поколение эпигеномное влияние может сформироваться, например, под действием вируса, инфицирующего исходную и попадающего в каждую новообразовавшуюся клетку в митозе (рис. 13.3). Допускается, что в геноме вируса имеется ген типа клеточного гена-репрессора. 2. C появлением вируса в клетке начинается синтез вирусного репрессора 2, который тормозит работу гена-репрессора 1, и репрессор 1 не синтезируется. В отсутствие репрессора 1 активизируется . ген, кодирующий инициатор деления клетки, и синтезируется данный инициатор. Появление инициатора деления клетки приводит к началу репликации ДНК, имеющейся в геноме клетки, т. е. к делению клетки. При этом инициатор деления выключает ген-репрессор 2 клетки, но не может выключить онкогенную часть генома вируса - вирусный ген-репрессор типа клеточного гена-репрессора 2 в силу особенностей строения вирусного гена-репрессора 2.
Таким образом, пока вирус находится в клетке, постоянно поддерживается синтез репрессора 2 на основе вирусного гена. Это приводит к репрессии регуляторного гена-репрессора 1 клетки, в связи с чем дерепрессируется ген, кодирующий инициатор деления клетки и клетка продолжает размножаться.Образовавшиеся клетки содержат в себе вирусный геном, который попадает к ним во время митоза от исходной клетки и поддерживает нарушение регуляции клеточного деления в последующих поколениях клеток. Получены доказательства, что онкогены опухолеродных вирусов действительно идентичны клеточным факторам роста. Более того, в клетках сетчатки найден ген-репрессор клеточного деления, названный Rb, наследственный дефект которого приводит у ребенка к развитию злокачественной опухоли сетчатки глаза - ретинобластомы.
Мутационный и эпигеномный механизмы канцерогенеза не исключают друг друга, а могут быть сопряженными. В клетке имеются специальные регуляторные гены, репрессирующие геном опухолеродного вируса (см. рис. 13.1). Следовательно, мутация может произойти с репрессорным геном клетки, в результате чего активируется вышедший из-под контроля онкоген опухолеродного вируса и происходит эпигеномная трансформация клетки. Таким образом, химические и физические факторы могут не сами по себе вызывать трансформацию, а способствовать активизации вирусного канцерогенеза.
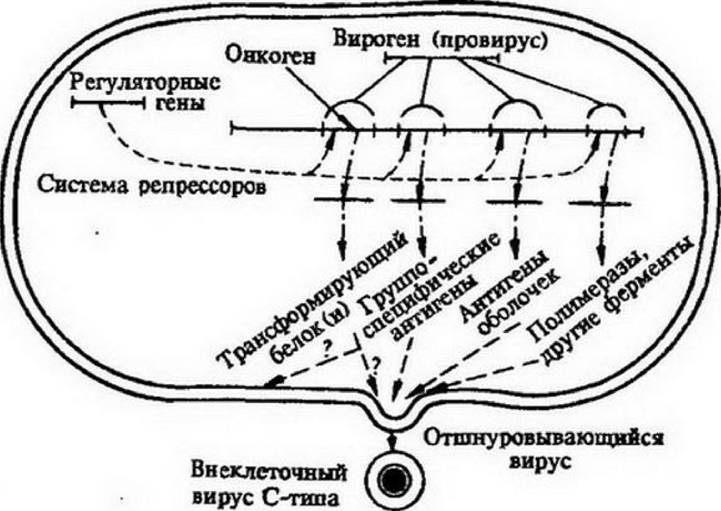
Рис. ІЗЛ. Модель контроля онко- и вирусогенеза.
Опыты Гердона по пересадке ядер дифференцированных клеток в цитоплазму зиготы были использованы для изучения регуляции деления в ядрах опухолевых клеток. Если из оплодотворенных яйцеклеток лягушки удаляют ядро и вместо него подсаживают ядро из специализированной клетки с двойным набором хромосом, например клетки кишечного эпителия, то развивается головастик со всеми дифференцирующимися органами. Следовательно, в ядрах специализированных клеток организма сохраняется весь набор генов, который был в ядре зиготы. Когда в цитоплазму зиготы подсадили ядро раковой клетки из почечной опухоли Люкке лягушки, также развился головастик, а не опухолевая ткань. После пересадки ядра клетки такого головастика в цитоплазму клетки опухоли Люкке образовалась нормальная почечная ткань.
Таким образом, здоровая цитоплазма зиготы обусловила полное восстановление регуляции в пересаженном ядре опухолевой клетки и закрепление его как наследуемое свойство. Поскольку опухоль Люкке лягушек имеет вирусное происхождение, следовательно, речь идет о восстановлении функции генов, репрессирующих вирусный геном.
Еще по теме Мутационный канцерогенез.:
- К А Н Ц Е Р О Г Е Н Е З
- РОЛЬ НАСЛЕДСТВЕННЫХ ФАКТОРОВ В ЭТИОЛОГИИ И ПАТОГЕНЕЗЕ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ
- БИОЛОГИЧЕСКАЯ СУЩНОСТЬ (ПРИРОДА) ОПУХОЛЕВОГО РОСТА
- Оглавление
- Патогенез
- Мутационный канцерогенез.
- ТРАНСФОРМАЦИЯ ЗДОРОВОЙ КЛЕТКИ В ОПУХОЛЕВУЮ
- Современные представления об этиопатогенезе злокачественных новообразований яичников